Docosahexaenoic acid (DHA) is a long-chain omega-3 (n-3) polyunsaturated fatty acid that is a critical component of lipid structures. DHA plays important roles throughout the body and is essential for maintaining the structure and function of the brain and eye. Fetal development and infancy are key windows during which sufficient DHA levels are necessary for optimal mental and visual development and performance in later life.
Current knowledge
Humans have a limited ability to synthesise DHA from essential fatty acids; the main source of DHA is from the diet. Transported in the blood as a component of lipoproteins, DHA can be stored in adipose tissues. Compared to other bodily tissues, the eye and brain contain a high proportion of DHA. Due to its highly unsaturated composition, DHA adopts a three-dimensional shape that is different from that of other fatty acids in cell membranes. In the rod cells of retinal photoreceptors for example, DHA within the membrane facilitates the conformational change triggered by a light signal. In addition to its effects in the eye and brain, DHA also reduces inflammation, improves immune function, and optimises cellular metabolism.
Practical implications
Maternal plasma phospholipids are an important source of DHA for the fetus. Indeed, DHA is highly concentrated in the fetal circulation and in fetal tissues through the process of biomagnification. DHA is naturally found in breast milk. The DHA content of breast milk can be increased by maternal consumption of DHA-rich foods such as fish, eggs, fish oil, or DHA-rich oil. Higher consumption of DHA by lactating women results in increased DHA in breast milk, ultimately raising the infant’s DHA status. Therefore, ensuring sufficient dietary DHA in pregnant women is key for optimal fetal development.
Recommended reading
Childs CE, Romeu-Nadal M, Burdge GC, Calder PC: Gender differences in the n-3 fatty acid content of tissues. Proc Nutr Soc 2008;67:19–27
Docosahexaenoic acid (DHA) is a bioactive omega-3 polyunsaturated fatty acid that influences membrane structure and function, cell signalling and communication mechanisms, gene expression and lipid mediator production.
DHA is found in high concentrations in the human brain and eye, where it is linked to better development and function.
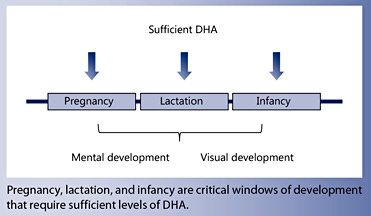
Maintenance of DHA concentration is important throughout the life course, but pregnancy, lactation and infancy are vulnerable periods, where insufficient DHA supply can impact mental and visual development and performance.
Key Words
Docosahexaenoic acid · Fish · Fish oil · Omega-3 · Pregnancy · Lactation · Life course
Abstract
Docosahexaenoic acid (DHA) is a long-chain, highly unsaturated omega-3 (n-3) fatty acid. It has a structure that gives it unique physical and functional properties. DHA is metabolically related to other n-3 fatty acids: it can be synthesised from the plant essential fatty acid α-linolenic acid (ALA). However, this pathway does not appear to be very efficient in many individuals, although the conversion of ALA to DHA is much better in young women than in young men. Furthermore, young infants may be more efficient converters of ALA to DHA than many adults, although the conversion rate is variable among infants. Many factors have been identified that affect the rate of conversion. The implication of poor conversion is that preformed DHA needs to be consumed. DHA is found in fairly high amounts in seafood, especially fatty fish, and in various forms of n-3 supplements. The amount of DHA in seafood and in supplements varies. Breast milk contains DHA. DHA is found esterified into complex lipids within the bloodstream, in adipose stores and in cell membranes. Its concentration in different compartments varies greatly. The brain and eye have high DHA contents compared to other organs. DHA is especially concentrated in the grey matter of the brain and in the rod outer segments of the retina. In the brain DHA is involved in neuronal signalling, while in the eye it is involved in the quality of vision. DHA is accumulated in the brain and eye late in pregnancy and in early infancy. A lower DHA content is linked to poorer cognitive development and visual function. DHA affects cell and tissue physiology and function through numerous mechanisms, including alterations in membrane structure and function, in membrane protein function, in cellular signalling and in lipid mediator production.
Introduction
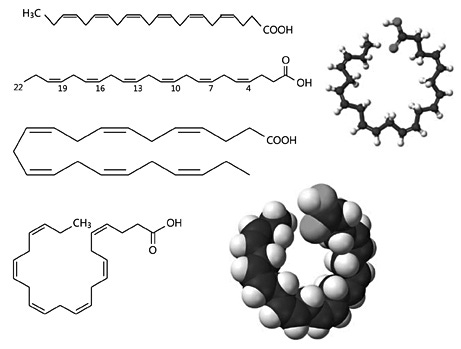
Docosahexaenoic acid (DHA) is a long-chain, highly unsaturated omega-3 (n-3) fatty acid (see Appendix). It has 22 carbons in its acyl chain, which includes 6 double bonds. Chemically it can be described as all- cis - 4,7,10,13,16,19-docosahexaenoic acid, with the numbers 4, 7, 10, 13, 16 and 19 referring to the carbon atoms in the acyl chain that bear double bonds when the carboxyl or α-carbon is counted as number 1 ( fig. 1 ). DHA is shown in the common fatty acid nomenclature as 22: 6ω-3 or 22: 6n-3, with the ω-3 (or n-3) indicating the position of the first double bond in the acyl chain, in this case counting the methyl or ω-carbon as number 1 (fig. 1). The common name for DHA, which is rarely used, is cervonic acid. Fatty acyl chains with no double bonds, such as in saturated fatty acids, are straight and pack together tightly. Introduction of a cis double bond into an acyl chain introduces a ‘kink’ into the chain, making it less easy for such chains to pack together and lowering their melting point. As the acyl chain of DHA contains 6 cis double bonds, it becomes highly twisted (fig. 1), giving it unique physical properties and resulting in a very low melting point (–44° C). DHA is metabolically related to other n-3 fatty acids. It can be synthesised from the plant-derived α-linolenic acid (ALA; 18: 3n-3) or obtained directly from the diet. In common with other fatty acids, DHA is most often found linked via its carboxyl group into a more complex lipid structure such as a triglyceride, phospholipid or cholesteryl ester. Here, the pathway of DHA biosynthesis, dietary sources of DHA, the status (i.e. concentration) of DHA at different sites in the human body and the response of those sites to increased DHA intake, and selected actions of DHA at the molecular and cellular levels will be described. DHA plays vital roles in the structure and function of the brain and eye and an appropriate supply during fetal life and in infancy is essential to assure optimal development.
Fig. 1. Different depictions of the structure of DHA. DHA has 22 carbons and 6 cis double bonds in its hydrocarbon (acyl) chain. The α-carbon is the carbon of the terminal carboxyl group (COOH) and the ω-carbon is the carbon of the terminal methyl (CH 3 ) group.
Biosynthesis of DHA
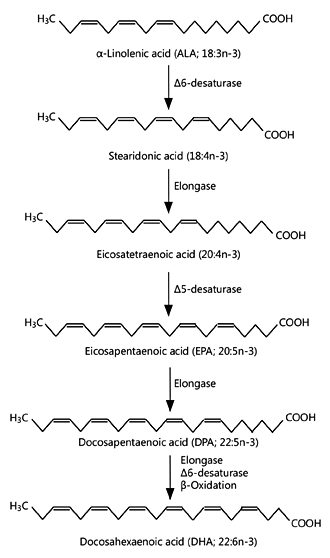
ALA is an essential fatty acid. It is synthesised in plants and in many lower organisms and is found in the human diet mainly as a component of green leaves, some nuts, seeds and vegetable oils, and foods made from or containing those ingredients. There is a metabolic pathway that links ALA to DHA (fig. 2). This pathway involves a series of enzyme-catalysed elongation and desaturation reactions. Elongation enzymes, called elongases, add pairs of carbon atoms to the growing acyl chain, in this case converting an 18-carbon fatty acid into a 22-carbon one, while desaturase enzymes insert double bonds into the acyl chain, in this case converting a fatty acid with 3 double bonds in its acyl chain into one with 6 double bonds. These reactions occur predominantly within the endoplasmic reticulum. The pathway is believed to mainly occur within the liver, but there is some evidence that other tissues, including brain and testis, have high expression of the genes encoding the relevant enzymes.
The initial step in the pathway is the conversion of ALA to stearidonic acid (18: 4n-3), catalysed by Δ-6- desaturase, which is generally considered to be the ratelimiting reaction in the pathway. Δ-6-desaturase is encoded by the gene fatty acid desaturase 2 (Fads2) . Stearidonic acid is converted to 20: 4n-3 by the addition of 2 carbons by the enzyme elongase-5, encoded by fatty acid elongase 5 (Elovl5). 20: 4n-3 is then converted to eicosapentaenoic acid (EPA; 20: 5n-3) by insertion of a double bond catalysed by Δ-5-desaturase, which is encoded by the gene fatty acid desaturase 1 (Fads1) . EPA can be elongated by elongase 2 (encoded by Elovl2 ) to form n-3 docosapentaenoic acid (DPA; 22: 5n-3) and then to 24: 5n-3 followed by desaturation that again uses Δ-6-desaturase activity to form 24: 6n-3. This desaturation seems to be catalysed by the same Δ-6-desaturase as in the first step of the pathway. 24: 5n-3 is then translocated from the endoplasmic reticulum to the peroxisome where it undergoes one round of β-oxidation to form DHA.
It is important to note that the same enzymes are active in the metabolism of the n-6 fatty acid family, converting the essential n-6 fatty acid linoleic acid (18: 2n-6) to arachidonic acid (20: 4n-6) and on to n-6 DPA (22: 5n-6). Thus, competition exists between the conversion of n-6 and n-3 fatty acids. The rate-limiting enzyme, Δ-6- desaturase, has a preference for ALA over linoleic acid. However, this may be more than offset by the greater abundance of linoleic acid than ALA in most human diets, meaning that the metabolism of the former is favoured.
In addition to the availability of the essential fatty acid substrates and the competition between them, a number of other factors have been demonstrated to regulate the pathway. These include the availability of several trace elements including zinc and iron, since the enzymes involved in the pathway require these as co-factors; sensitivity to insulin; female sex hormone status; polymorphisms in Fads genes which control gene expression and enzyme activity, and epigenetic modification of Fads and Elovl genes, which will affect their expression. Other nutrients, metabolites and hormones, and ageing may also affect the pathway. Measurements of EPA and DHA status reveal differences among some population subgroups, for example between men and women [1, 2] and among individuals with different Fads polymorphisms [3], that are likely to reflect different activities of the biosynthetic pathway. There has been much interest in the reported differences in EPA and DHA status between men and women. Studies using stable isotopes to trace metabolism of ALA have demonstrated that the conversion of ALA to both EPA and DHA is more efficient in young women than in young men [4, 5]. In men, conversion of ALA to EPA has been reported to be between 0.3 and 8%, and conversion to DHA <1%, whereas in women up to 21% conversion to EPA and up to 9% conversion to DHA have been reported. It has been suggested that the higher rate of conversion in women may be because of their greater requirement to produce DHA during pregnancy and lactation. Infants may be more effective at converting ALA to DHA than adults, and newborns appear to be better at synthesising DHA than older infants [6].
Fig. 2. The metabolic pathway of conversion of α-linolenic acid to DHA showing the enzymes involved.
Dietary Sources of DHA
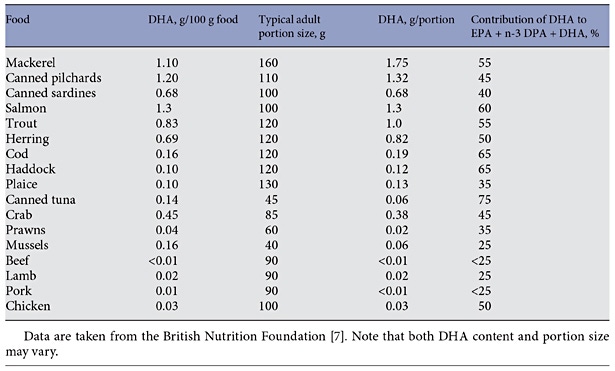
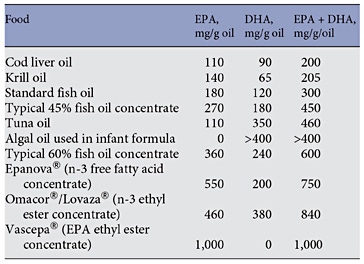
Along with EPA and n-3 DPA, DHA is found in fairly high amounts in seafood and products derived from seafood. Table 1 reports typical values for the content of DHA in various seafoods [7]. It is evident that there is at least a 10-fold range in content of DHA per portion (i.e. per serving) of seafood, with fatty fish able to provide as much as 1–1.75 g of DHA per portion. Examples of fatty fish are mackerel, salmon, trout, herring, tuna and sardines. In comparison, lean fish like cod, haddock and plaice typically provide about 0.1–0.2 g of DHA per portion. Although tuna is a fatty fish, canned tuna has had the oil removed during processing and so is low in content of n-3 fatty acids including DHA (table 1). Meat and blubber of sea mammals like seals and whales is also rich in n-3 fatty acids including DHA, although these are not usually eaten by most humans. Likewise, some organ meats like brain are rich in DHA but again these are rarely eaten in most populations. DHA is found in modest amounts in animal-derived foods like eggs and meat (table 1). Human breast milk and other mammalian milks contain DHA. An analysis of data from 65 studies of over 2,400 women from around the globe gave a mean concentration of DHA in breast milk as 0.32% of total fatty acids by weight with a range of 0.06–1.4% [8]. The highest breast milk DHA concentrations were found in coastal populations and were associated with marine food consumption [8].
Because fatty fish are the richest dietary source of DHA, DHA intake is heavily influenced by fish consumption. In most Western populations, the distribution of fatty fish consumption is bimodal, with a relatively small proportion of the population (e.g. 15–25%) being regular fatty fish consumers. Thus, typical intakes of DHA across a population are likely to be low. Data from over 10,000 Australian adults identified a mean daily DHA intake of 106 mg, with DHA contributing about 60% of n-3 longchain polyunsaturated fatty acid (LCPUFA) intake [9]. By comparison, mean daily intakes of EPA and n-3 DPA were 56 and 25 mg, respectively [9]. However, the median intake of DHA was only 15 mg/day [9], the large differences between mean and median intakes reflecting the non-normal distribution of the intake data. Median daily intakes of EPA and n-3 DPA were 8 and 6 mg, respectively [9]. In a more recent study using an updated nutrient composition database, the mean daily intake for DHA was 100 mg, which was 40% of n-3 LCPUFA intake [10]. Again, median intakes were lower, being about 50% of the mean. An interesting observation from these studies is that meat (including poultry) provides 40–45% of EPA + n-3 DPA + DHA consumed by typical Australian adults, with fish and seafood products providing 45–50% [10].
DHA is also present in fish body (aka fish), fish liver, algal and krill oils, in concentrated supplements and in pharmaceutical grade preparations designed for controlling blood triglyceride levels. Table 2 summarises the DHA content of typical preparations of these sources. It is clear that the use of supplements, whether these are over-the-counter or prescribed, can substantially increase the intake of DHA. DHA-rich oils from algae or tuna are used in infant formulas.
Table. 1. Typical DHA content of selected fish, other seafood and meats
Table. 2. Typical EPA and DHA contents of fish oils and other n-3 supplements
DHA Status
DHA Concentration in Different Metabolic and Anatomical Compartments
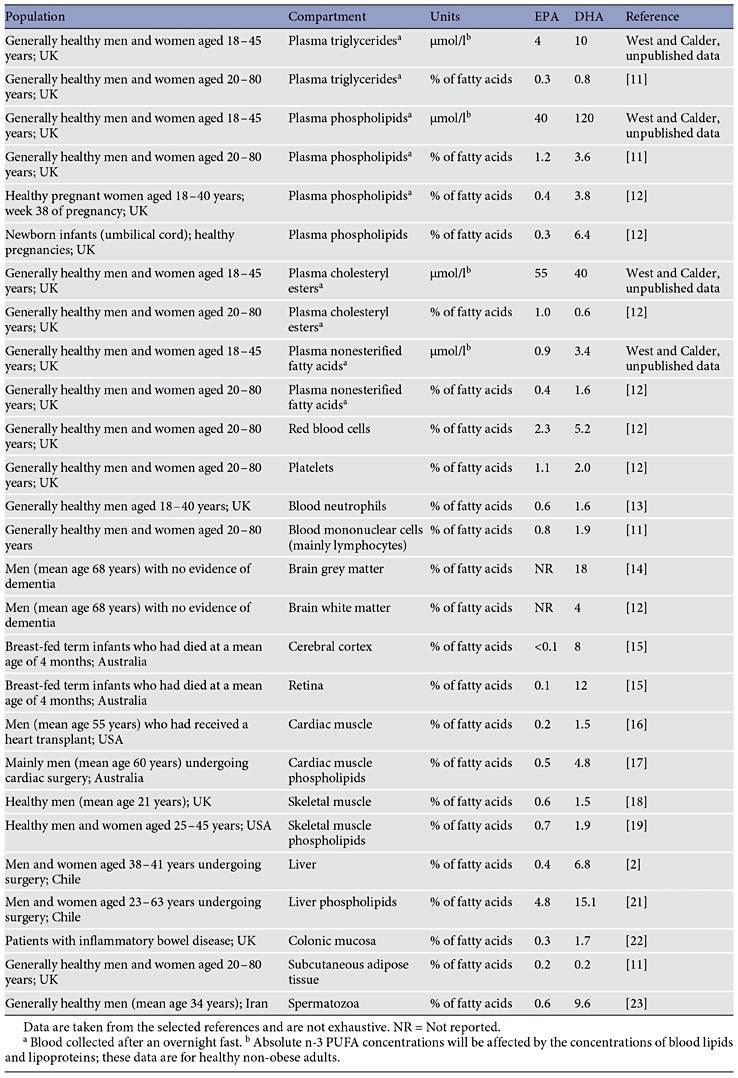
DHA is transported in the bloodstream as a component of lipoproteins (within triglycerides, phospholipids and cholesteryl esters) or as a nonesterified (‘free’) fatty acid (largely due to release from adipose tissue stores or from spill-over of lipase-mediated hydrolysis of circulating lipoproteins). DHA can be stored in adipose tissue esterified into triglycerides. DHA is found in all cell membranes esterified into phospholipids and other complex lipids. Table 3 lists some reported concentrations of EPA and DHA in different metabolic or anatomical compartments in humans [11–23] . It is evident that the absolute and proportional contribution of EPA or DHA to the total fatty acids present within any of the compartments listed in table 3 differs both between the two fatty acids and between compartments. Most often, DHA is present at a higher concentration than EPA. This is especially true in specific regions of the brain and eye, where DHA makes a significant contribution to the fatty acid complement and EPA is virtually absent.
DHA is Highly Concentrated in the Human Brain and Eyes
More than 50% of the dry weight of the human brain is lipid, particularly structural lipid (i.e. phospholipids). The most abundant fatty acids in the brain are DHA, arachidonic acid and adrenic acid [24, 25]. The human brain and retina contain an especially high proportion of DHA relative to other tissues and little EPA [24–26] (table 3). For example, DHA was reported to contribute an average of 18% of fatty acids to adult human brain grey matter [14], while Makrides et al. [15] reported average DHA contents of about 8 and 12% of fatty acids for human infant cerebral cortex and retina, respectively. In the latter study, the contributions of EPA were <0.05 and 0.1%, respectively [15]. Within cell membranes, EPA and DHA are distributed differently among the different phospholipid components and in the brain and eye specific phospholipids are especially rich in DHA. For example, DHA was reported to contribute an average of 36% of fatty acids in mammalian brain grey matter phosphatidylserine [25] and an average of 22% of fatty acids in retina phosphatidylcholine [26]. DHA contributes 50–70% of the fatty acids present in the rod outer segments of the retina [26]. These rod outer segments contain the eyes’ photoreceptors.
The human brain growth spurt occurs from approximately the beginning of the third trimester of pregnancy to 18 months after birth. The amount of DHA in the brain increases dramatically during the brain growth spurt. In humans, brain weight increases from about 100 g at 30 weeks of gestation to about 1,100 g at 18 months of age [27]. Over this period, the DHA content of the brain increases from 900 μg/g (90 mg in total) to 3,000 μg/g (3,300 mg total) [28, 29]. This represents a 35-fold increase in total brain DHA. The estimated rate of accretion of DHA into the human brain in the last trimester of pregnancy is 15–22 mg/week [30]. This is also the most active period of brain cell division. Thus, it is believed that an adequate supply of DHA during this period is essential for normal growth, neurological and visual development and function, and learning behaviour [29].
DHA Concentration during Pregnancy
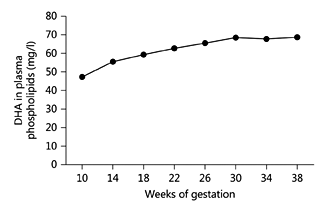
There is a significant linear relationship between the DHA contents of maternal and umbilical cord plasma phospholipids. This suggests that maternal plasma phospholipids are an important source of DHA for the fetus and that maternal plasma phospholipid DHA concentration determines DHA supply to the fetus. An increase in maternal plasma DHA concentration occurs during pregnancy [31] (fig. 3), and this increase precedes the increase in DHA accretion by the brain. The extent of the increase differs among pregnant women in different countries [32], perhaps indicating dietary differences during the pre-pregnancy period. Al et al. [33] reported that maternal plasma phospholipid DHA content was lower in women who had had multiple pregnancies than in those in their first pregnancy. This may indicate that maternal body stores are important in maintaining plasma DHA status but that these may be eroded by multiple pregnancies.
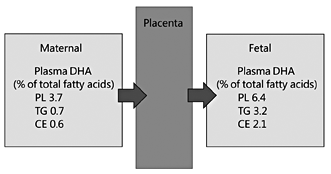
Even though maternal and fetal blood DHA concentrations are highly correlated, DHA is concentrated in the fetal circulation and in fetal tissues [34], a process sometimes referred to as biomagnification (fig. 4). Placental adaptation to ensure efficient DHA transfer from the maternal to the fetal circulation is an important part of the biomagnification process; this is discussed in detail elsewhere [34]. Furthermore, there are observations that the placenta can synthesise DHA from ALA [34], which would allow for in situ provision of DHA to help meet the demands for DHA imposed by pregnancy.
Table 3. Typical EPA and DHA concentrations reported in different compartments in humans
Fig. 3. The change in DHA concentration (mg/l) in plasma phospholipids during human pregnancy. Maternal blood was collected from the same women at different time points in pregnancy. Data are taken from Al et al. [31] and represent the mean values from 110 women.
Fig. 4. DHA concentration (as % of total fatty acids) in maternal blood plasma lipids in late pregnancy and in umbilical cord blood plasma at birth. Maternal blood was collected at week 38 of pregnancy. Data are previously unpublished and represent mean values from 50 pregnancies. PL = Phospholipid; TG = triglyceride; CE = cholesteryl ester.
The Effect of Increased DHA Intake on DHA Status
Increasing the Intake of DHA Increases Its Status in Many Body Compartments
Increased intakes of DHA (and EPA) from fish or from n-3 supplements are reflected in increased proportions of both fatty acids in blood lipid, blood cell and many tissue compartments. This has been reported many times for total plasma or serum lipids or for the complex lipid components of plasma or serum (i.e. triglycerides, phospholipids and cholesteryl esters) and is also well described for erythrocytes, platelets and leukocytes (see [35] for references). There are also descriptions of increased proportions of DHA (and EPA) in human tissues, including skeletal muscle [18, 19], heart [16], gut mucosa [22] and adipose tissue [11, 36] when their intake is increased. These compartments all show a dose- and time-dependent incorporation of both EPA and DHA [11, 13, 35, 36] , but the precise pattern depends upon the specific location [37, 38] (fig. 5). Pools that are turning over rapidly show faster incorporation of both EPA and DHA than slower turning-over pools. Thus, plasma lipids incorporate EPA and DHA more quickly than blood cells do [11, 36], whilst amongst blood cells, leukocytes have been usually shown to incorporate EPA and DHA more quickly than erythrocytes. Modification of human brain fatty acid composition is more difficult than for other tissues, especially beyond childhood. However, Makrides et al. [15] demonstrated that the DHA concentration of the cerebral cortex of breast-fed infants who would have received DHA via breast milk is higher than that of infants who had not been breast fed.
The higher status of EPA and DHA achieved through increased intake is maintained so long as the higher intake of EPA and DHA is maintained. If, after a period of increased intake of EPA and DHA, the intake returns to the earlier lower levels, then EPA and DHA statuses decline, eventually returning to earlier levels (fig. 5). This is well described for blood lipids, platelets, leukocytes and erythrocytes [35]. However, just as the incorporation of EPA into different pools is faster than the incorporation of DHA, the loss of EPA is faster than the loss of DHA [35]. One interpretation of this preferential retention of DHA is that DHA is structurally and/or functionally preferred over EPA and that metabolic mechanisms have evolved to preserve it.
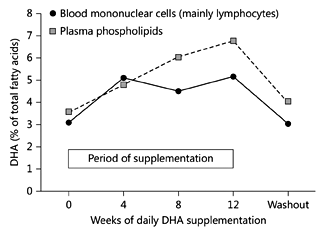
Fig. 5. DHA concentration (as % of total fatty acids) in plasma phospholipids and peripheral blood mononuclear cells from older subjects supplemented with DHA (720 mg/day) for 12 weeks. Blood was collected at baseline and after 4, 8 and 12 weeks of supplementation with near pure DHA (algal source) and again 4 weeks after stopping supplementation. Data are taken from Thies et al. [37, 38] and represent mean values from 8 healthy older subjects (age 58–71 years).
Increasing Maternal and Fetal DHA Status during Pregnancy
With the recognition that maternal supply of DHA to the fetus is important and that fetal and infant DHA status is a major contributor to optimal visual and brain development and function, investigations of ways to increase maternal DHA status have been made. Clearly the simplest way to do this would be to increase maternal DHA intake from fish or from supplements. Regular consumption of fatty fish resulted in higher maternal erythrocyte DHA content [39, 40]. Connor et al. [41] reported that consumption of sardines and fish oil by women from week 30 of pregnancy resulted in higher DHA contents of maternal plasma and erythrocytes 4 weeks later than observed in women taking placebo. In that study, a total of 3 g n-3 LCPUFAs including 1.1 g DHA were supplied per day. A similar intake of n-3 LCPUFAs (2.7 g/day) and of DHA (1.1 g/day) provided as fish oil from week 30 of pregnancy resulted in higher DHA contents of maternal plasma phospholipids and of umbilical cord vein and artery at birth [42]. Fish oil providing 3.3 g/day n-3 LCPUFAs (including 2.2 g/day as DHA) from week 20 of pregnancy resulted in higher DHA contents of maternal erythrocytes at weeks 30 and 37 of pregnancy and at 6 weeks after delivery [43]. Furthermore, the DHA content of cord blood erythrocytes was 40% higher than in the placebo group [43]. A much lower intake of DHA (200 mg/day) from week 15 of pregnancy resulted in higher DHA content of maternal plasma erythrocytes at week 28 of pregnancy and at birth, although there was no effect on DHA content of cord blood plasma or erythrocytes [44].
Increasing the Status of DHA in Human Breast Milk
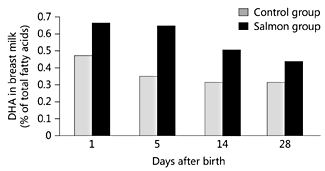
After a baby is born it is important that a supply of preformed DHA is continued. Breast milk DHA content can be increased by maternal supplementation with fish oil [45], DHA-rich oil [46, 47] or n-3 LCPUFA-rich eggs [47] or by increased maternal fatty fish consumption [48] . Harris et al. [45] gave lactating women 5 g/day fish oil for 28 days, 10 g/day fish oil for 14 days or 47 g/day fish oil for 8 days. Levels of DHA in breast milk were 0.1% of total fatty acids at baseline and 0.5, 0.8 and 4.8% of fatty acids after 5, 10 and 47 g/day fish oil, respectively. Hawkes et al. [46] gave lactating women placebo, ‘low’-dose DHA (300 mg/day) or ‘high’-dose DHA (600 mg/day) from day 3 after delivery for 4 weeks. They found that the DHA content of maternal plasma, maternal mononuclear cells, breast milk and breast milk cells increased in relation to DHA intake. In another low-dose approach, Jensen et al. [47] compared fish oil, algal oil and eggs as a source of DHA for incorporation into breast milk: lactating women (2 weeks after giving birth) received about 200 mg/day DHA from the oils or eggs for 4 weeks. This resulted in an increase in DHA in maternal plasma phospholipids (from 2.5% to about 4% of total fatty acids) and in breast milk (from about 0.2% to about 0.4% of total fatty acids). Furthermore, infant plasma phospholipid DHA increased from about 3.6% to about 5% of total fatty acids. This demonstrates that increased consumption of DHA by lactating women results in increased DHA in breast milk, subsequently elevating infant DHA status. The Salmon in Pregnancy Study investigated the effect of consumption of two portions of salmon per week from week 20 of pregnancy until delivery [12]. This intervention resulted in higher breast milk DHA than seen in women in the control group who did not eat fatty fish [48] (fig. 6).
Fig. 6. The effect of eating salmon during pregnancy on DHA concentration (as % of total fatty acids) in human breast milk. Pregnant women consumed two portions of salmon per week from week 20 of pregnancy until birth. The control group reported very low or no consumption of fatty fish. Breast milk was collected at days 1, 5, 14 and 28 after giving birth. Data are taken from Urwin et al. [48] and represent mean values from 7 to 27 samples depending upon group and time point.
Mechanisms of Action of DHA
Cell Membranes Containing DHA within Phospholipids Have Unique Functional Properties
Phospholipids are quantitatively the major lipid component of cell membranes. Fatty acids in the phospholipids play important roles assuring the correct environment for membrane protein function, maintaining membrane order (‘fluidity’), and influencing the formation of signalling platforms termed lipid rafts. Membrane phospholipids are substrates for the generation of second messengers like diacylglycerols, lysophospholipids and PUFAs themselves [49]. DHA is a constituent of cell membrane phospholipids and imparts unique physical and chemical properties on the phospholipid and any signalling molecules that are produced from it [49]. Because of its highly unsaturated nature, DHA adopts a three-dimensional shape that is different from that of other common membrane fatty acids (fig. 1). This shape strongly influences membrane order and has an impact on membrane protein function and on the assembly of lipid rafts (fig. 7). Hence, the DHA content of a cell membrane can have a significant influence on cellular behaviour and responsiveness to signals, which may be electrical, chemical, hormonal or antigenic in nature. Within the brain, DHA has important actions in regulating intracellular signalling [50]. Perhaps the single best example of the unique role that cell membrane DHA has in a physiological function relates to the role of the rod in the retinal photoreceptor [51]. The cells of the rod outer segment have an exceptionally high content of DHA in their membranes (50–70% of fatty acids) [26]. The DHA is a component of phospholipids that cluster around the protein rhodopsin which receives the light signal. When the signal is received, rhodopsin undergoes a conformational change that initiates a signal transduction cascade. The physical nature of DHA within the membrane facilitates the conformational change [51]. Studies with rhodopsin imbedded in artificial membrane bilayers demonstrate that phospholipids containing two DHA molecules are greatly superior to phospholipids containing one or no DHA molecules, even when they contain other highly unsaturated fatty acids such as n-6 DPA or arachidonic acid [52].
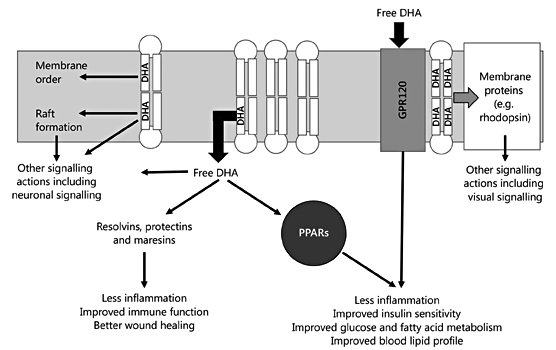
Fig. 7. A schematic summary of the mechanisms of action of DHA.
DHA Can Act via Cell Surface and Intracellular Receptors
Peroxisome Proliferator-Activated Receptors Peroxisome proliferator-activated receptors (PPARs) are transcription factors; they regulate gene expression and so play a role in cell and tissue responses to the environment. There are several isoforms of PPARs; PPAR-α and PPAR-γ are the most well understood. PPARs act by forming a heterodimer with the retinoic X receptor, the ligand for which is cis-9-retinoic acid. PPAR-α is expressed mainly in the liver and is involved in regulating hepatic metabolic responses. The transcription of genes encoding several key enzymes of β-oxidation and of lipoprotein metabolism has been shown to be regulated by PPAR-α. Thus, activation of PPAR-α results in partitioning of fatty acids towards hepatic oxidation and away from triglyceride synthesis. PPAR-γ is expressed in adipose tissue, where it is involved in regulating adipocyte differentiation and metabolic responses of adipocytes including promoting insulin sensitivity, and in inflammatory cells, where it is involved in regulating the production of inflammatory mediators, having an anti-inflammatory action. PPARs are activated by noncovalent binding of ligands which include n-3 LCPUFAs and various lipid mediators. DHA can induce and activate PPARs [53] and upregulate a number of PPAR target genes [54], which may be at least partly responsible for the ability of DHA to lower fasting plasma triglyceride concentrations [55], to increase insulin sensitivity [56] and to reduce inflammation [57].
G-Protein-Coupled Receptors Several membrane-associated G-protein-coupled receptors (GPRs) are able to bind fatty acids differentially according to structural features of those fatty acids. GPR40 and GPR120 can both bind long-chain fatty acids and are active in signal transduction. Oh et al. [58] reported that GPR120 is highly expressed on adipocytes and on inflammatory macrophages. Macrophage GPR120 was shown to be involved in anti-inflammatory signalling, inhibiting activation of the prototypical pro-inflammatory transcription factor nuclear factor kappa B [58]. DHA (and EPA) promoted GPR120-mediated gene activation, and several anti-inflammatory actions of DHA did not
occur in GPR120 knockdown cells. Oh et al. [58] also demonstrated that DHA promoted the translocation of the glucose transporter GLUT4 to the surface of cultured adipocytes and that this was associated with enhanced glucose uptake. These effects were abolished by GPR120 knockout, suggesting that GPR120 mediates at least some of the metabolic actions of DHA.
DHA is a Substrate for Biosynthesis of Bioactive Mediators
PUFAs including linoleic acid, dihomo-γ-linolenic acid (20: 3n-6), arachidonic acid, EPA, n-3 DPA, n-6 DPA (22: 5n-6) and DHA all give rise to bioactive lipid mediators formed through the cyclooxygenase and lipoxygenase pathways [57]. These mediators have many actions, but are most well recognised for their roles in inflammation, immunity, platelet reactivity and smooth muscle contraction. Historically, most attention has been given to the prostaglandins, thromboxanes and leukotrienes, produced from arachidonic acid. Like EPA, DHA has some actions to reduce the production of these mediators, which may be an important mechanism by which DHA can affect inflammation, immunity, blood clotting and so on. More importantly, however, new families of bioactive lipid mediators produced from both EPA and DHA have been discovered. These mediators are able to induce resolution of inflammation (i.e. ‘turn inflammation off’) and to promote immune function, thereby assisting with host defence and diminishing the pathological effect of inflammation [59, 60]. They include the resolvins produced from EPA (E-series) and DHA (D-series) and protectins and maresins produced from DHA. Protectins are also referred to as neuroprotectins when generated within neural tissue, where they appear to have an important role [59, 60]. The synthesis of resolvins, protectins and maresins involves the cyclooxygenase and lipoxygenase pathways, with different epimers being produced in the presence and absence of aspirin [59, 60] . Figure 7 shows the outline of the pathway of DHA conversion to these mediators. D-series resolvins and protectins have been described in human plasma [61], adipose tissue [62] and breast milk [63]. The biological effects of resolvins, protectins and maresins have been examined extensively in cell culture and in animal models [59, 60]. Resolvins D1 and D2 and protectin D1 have each been demonstrated to have potent anti-inflammatory and inflammationresolving actions in these model systems [59, 60], acting through specific GPRs.
Conclusions
DHA is a long-chain, highly unsaturated n-3 fatty acid. It has a structure that gives it unique physical and functional properties. DHA is metabolically related to other n-3 fatty acids: it can be synthesised via EPA from the plant essential fatty acid ALA. However, this pathway does not appear to be very efficient in many individuals, although the conversion of ALA to DHA is much better in young women than in young men. It also appears that young infants may be more efficient converters of ALA to DHA than many adults, although the conversion rate is variable among infants. Many factors have been identified that affect the rate of conversion of ALA to DHA. These factors include n-6 fatty acid levels, availability of several trace elements, sensitivity to insulin, female sex hormone status and polymorphisms in, and epigenetic modification of, the genes encoding the various enzymes involved in the pathway. The implication of poor conversion is that DHA needs to be consumed preformed. Along with EPA and n-3 DPA, DHA is found in fairly high amounts in seafood, especially fatty fish, and in various forms of n-3 supplements. The amount of DHA in seafood and in supplements varies. Breast milk contains DHA. DHA is found esterified into complex lipids within the bloodstream, in adipose stores and in cell membranes. Its concentration in different compartments varies greatly. The brain and eye have high DHA contents compared to other organs. DHA is especially concentrated in the grey matter of the brain and in the rod outer segments of the retina. In the brain DHA is involved in neuronal signalling, while in the eye it is involved in visual quality. DHA is accumulated in the brain and eye late in pregnancy and in early infancy. A lower DHA content is linked to poorer cognitive development and visual function. Thus, it is vital that pregnant and lactating women as well as infants consume sufficient preformed DHA to support brain and eye development and function.
It is vital that pregnant and lactating women as well as infants consume sufficient preformed DHA to support brain and eye development and function
Increased intake of DHA by pregnant women and by infants results in higher DHA concentration in most body compartments. Breast milk DHA concentration is strongly influenced by dietary DHA intake. DHA affects cell and tissue physiology and function through numerous mechanisms, which are summarised in figure 7. These include alterations in membrane structure and function, in membrane protein function, in cellular signalling and in lipid mediator production. In addition to the effects on neuronal signalling and vision, DHA reduces inflammation, improves immune function and optimises cellular metabolism. Through these effects DHA acts to lower the risk of insulin resistance, metabolic syndrome, hyperlipidemia and cardiovascular disease.
Disclosure Statement
P.C.C. is a consultant/advisor to Danone Nutricia Research, Pronova BioPharma, DSM, Cargill, Smartfish, Fresenius Kabi, Baxter Healthcare and Nestlé. The writing of this article was supported by Nestlé Nutrition Institute.
References
Childs CE, Romeu-Nadal M, Burdge GC, Calder PC: Gender differences in the n-3 fatty acid content of tissues. Proc Nutr Soc 2008; 67: 19–27.
Lohner S, Fekete K, Marosvolgyi T, Decsi T: Gender differences in the long-chain polyunsaturated fatty acid status: systematic review of 51 publications. Ann Nutr Metab 2013; 62: 98–112.
Schaeffer L, Gohlke H, Muller M, Heid IM, Palmer LJ, Kompauer I, Demmelmair H, Illig T, Koletzko B, Heinrich J: Common genetic variants of the FADS1 FADS2 gene cluster and their reconstructed haplotypes are associated with the fatty acid composition in phospholipids. Hum Mol Genet 2006; 15: 1745–1756.
Burdge GC, Jones AE, Wootton SA: Eicosapentaenoic and docosapentaenoic acids are the principal products of alpha-linolenic acid metabolism in young men. Br J Nutr 2002; 88: 355–363.
Burdge GC, Wootton SA: Conversion of alpha- linolenic acid to eicosapentaenoic, docosapentaenoic and docosahexaenoic acids in young women. Br J Nutr 2002; 88: 411–420.
Carnielli VP, Simonato M, Verlato G, Luijendijk I, De Curtis M, Sauer PJ, Cogo PE: Synthesis of long-chain polyunsaturated fatty acids in preterm newborns fed formula with long-chain polyunsaturated fatty acids. Am J Clin Nutr 2007; 86: 1323–1330.
British Nutrition Foundation: n-3 Fatty Acids and Health. London, British Nutrition Foundation, 1999.
Brenna JT, Varamini B, Jensen RG, Diersen- Schade DA, Boettcher JA, Arterburn LM: Docosahexaenoic and arachidonic acid concentrations in human breast milk worldwide. Am J Clin Nutr 2007; 85: 1457–1464.
Meyer BJ, Mann NJ, Lewis JL, Milligan GC, Sinclair AJ, Howe PR: Dietary intakes and food sources of omega-6 and omega-3 polyunsaturated fatty acids. Lipids 2003: 38; 391– 398.
Howe P, Meyer B, Record S, Baghurst K: Dietary intake of long-chain omega-311 polydounsaturated fatty acids: contribution of meat sources. Nutrition 2006; 22: 47–53.
Browning LM, Walker CG, Mander AP, West AL, Madden J, Gambell JM, Young S, Wang L, Jebb SA, Calder PC: Incorporation of eicosapentaenoic and docosahexaenoic acids into lipid pools when given as supplements providing doses equivalent to typical intakes of oily fish. Am J Clin Nutr 2012; 96: 748–758.
Miles EA, Noakes PS, Kremmyda LS, Vlachava M, Diaper ND, Rosenlund G, Urwin H, Yaqoob P, Rossary A, Farges MC, Vasson M-P, Liaset B, Frøyland L, Helmersson J, Basu S, Garcia E, Olza J, Mesa MD, Aguilera CM, Gil A, Robinson SM, Inskip HM, Godfrey KM, Calder PC: The Salmon in Pregnancy Study: study design, subject characteristics, maternal fish and marine n-3 fatty acid intake, and marine n-3 fatty acid status in maternal and umbilical cord blood. Am J Clin Nutr 2011; 94: 1986S– 1992S.
Healy DA, Wallace FA, Miles EA, Calder PC, Newsholme P: The effect of low to moderate amounts of dietary fish oil on neutrophil lipid composition and function. Lipids 2000; 35: 763–768.
Skinner ER, Watt C, Besson JA, Best PV: Differences in the fatty acid composition of the grey and white matter of different regions of the brains of patients with Alzheimer’s disease and control subjects. Brain 1993; 116: 717–725.
Makrides M, Neumann MA, Byard RW, Simmer K, Gibson RA: Fatty acid composition of brain, retina, and erythrocytes in breast- and formula-fed infants. Am J Clin Nutr 1994; 60: 189–194.
Harris WS, Sands SA, Windsor SL, Ali HA, Stevens TL, Magalski A, Porter CB, Borkon AM: Omega-3 fatty acids in cardiac biopsies from heart transplantation patients: correlation with erythrocyes and response to supplementation. Circulation 2004; 110: 1645– 1649.
Metcalf RG, James MJ, Gibson RA, Edwards JR, Stubberfield J, Stuklis R, Roberts-Thomson K, Young GD, Cleland LG: Effects of fish-oil supplementation on myocardial fatty acids in humans. Am J Clin Nutr 2007; 85: 1222–1228.
McGlory C, Galloway SD, Hamilton DL, Mc- Clintock C, Breen L, Dick JR, Bell JG, Tipton KD: Temporal changes in human skeletal muscle and blood lipid composition with fish oil supplementation. Prostaglandins Leukot Essent Fatty Acids 2014; 90: 199–206.
Smith GI, Atherton P, Reeds DN, et al: Omega- 3 polyunsaturated fatty acids augment the muscle protein anabolic response to hyperinsulinaemia- hyperaminoacidaemia in healthy young and middle-aged men and women. Clin Sci 2011; 121: 267–278.
Araya J, Rodrigo R, Videla LA, Thielemann L, Orellana M, Pettinelli P, Poniachik J: Increase in long-chain polyunsaturated fatty acid n-6/n-3 ratio in relation to hepatic steatosis in patients with non-alcoholic fatty liver disease. Clin Sci 2004; 106: 635–643.
Elizondo A, Araya J, Rodrigo R, Poniachik J, Csendes A, Maluenda F, Díaz JC, Signorini C, Sgherri C, Comporti M, Videla LA: Polyunsaturated fatty acid pattern in liver and erythrocyte phospholipids from obese patients. Obesity 2007; 15: 24–31.
Hillier K, Jewell R, Dorrell L, Smith CL: Incorporation of fatty acids from fish oil and olive oil into colonic mucosal lipids and effects upon eicosanoid synthesis in inflammatory bowel disease. Gut 1991; 32: 1151– 1155.
Safarinejad MR: Effect of omega-3 polyunsaturated fatty acid supplementation on semen profile and enzymatic anti-oxidant capacity of seminal plasma in infertile men with idiopathic oligoasthenoteratospermia: a double-blind, placebo-controlled, randomised study. Andrologia 2011; 43: 38–47.
O’Brien JS, Sampson EL: Fatty acid and fatty aldehyde composition of the major brain lipids in normal human gray matter, white matter, and myelin. J Lipid Res 1965; 6: 545–551.
Crawford MA, Casperd NM, Sinclair AJ: The long chain metabolites of linoleic avid linolenic acids in liver and brain in herbivores and carnivores. Comp Biochem Physiol B 1976; 54: 395–401.
Anderson RE: Lipids of ocular tissues. IV. A comparison of the phospholipids from the retina of six mammalian species. Exp Eye Res 1970; 10: 339–344.
Dobbing J, Sands J: Quantitative growth and development of human brain. Arch Dis Child 1973; 48: 757–767.
Cunnane SC, Francescutti V, Brenna JT, Crawford MA: Breast-fed infants achieve a higher rate of brain and whole body docosahexaenoate accumulation than formula-fed infants not consuming dietary docosahexaenoate. Lipids 2000; 35: 105–111.
Lauritzen L, Hansen HS, Jorgensen MH, Michaelsen KF: The essentiality of long chain n-3 fatty acids in relation to development and function of the brain and retina. Prog Lipid Res 2001; 40: 1–94.
Clandinin MT, Chappell JE, Leong S, Heim T, Swyer PR, Chance GW: Intrauterine fatty acid accretion rates in human brain: implications for fatty acid requirements. Early Hum Dev 1980; 4: 121–129.
Al MD, van Houwelingen AC, Kester AD, Hasaart TH, de Jong AE, Hornstra G: Maternal essential fatty acid patterns during normal pregnancy and their relationship to the neonatal essential fatty acid status. Br J Nutr 1995; 74: 55–68.
Otto SJ, Houwelingen AC, Antal M, Manninen A, Godfrey K, Lopez-Jaramillo P, Hornstra G: Maternal and neonatal essential fatty acid status in phospholipids: an international comparative study. Eur J Clin Nutr 1997; 51: 232–242.
Al MD, van Houwelingen AC, Hornstra G: Relation between birth order and the maternal and neonatal docosahexaenoic acid status. Eur J Clin Nutr 1997; 51: 548–553.
Haggarty P: Fatty acid supply to the human fetus. Ann Rev Nutr 2010; 30: 237–255.
Calder PC: Very long chain omega-3 (n-3) fatty acids and human health. Eur J Lipid Sci Technol 2014; 116: 1280–1300.
Katan MB, Deslypere JP, van Birgelen APJM, Penders M, Zegwaars M: Kinetics of the incorporation of dietary fatty acids into serum cholesteryl esters, erythrocyte membranes and adipose tissue: an 18 month controlled study. J Lipid Res 1997; 38: 2012–2022.
Thies F, Nebe-von-Caron G, Powell JR, Yaqoob P, Newsholme EA, Calder PC: Dietary supplementation with γ-linolenic acid or fish oil decreases T lymphocyte proliferation in healthy older humans. J Nutr 2001; 131: 1918–1927.
Thies F, Nebe-von-Caron G, Powell JR, Yaqoob P, Newsholme EA, Calder PC: Dietary supplementation with eicosapentaenoic acid, but not with other long chain n-3 or n-6 polyunsaturated fatty acids, decreases natural killer cell activity in healthy subjects aged >55 years. Am J Clin Nutr 2001; 73: 539– 548.
Olsen SF, Hansen HS, Sommer S, Jensen B, Sorensen TI, Secher NJ, Zachariassen P: Gestational age in relation to marine n-3 fatty acids in maternal erythrocytes: a study of women in the Faroe Islands and Denmark. Am J Obstet Gynecol 1991; 164: 1203–1209.
Sanjurjo P, Matorras R, Perteagudo L: Influence of fatty fish intake during pregnancy in the polyunsaturated fatty acids of erythrocyte phospholipids in the mother at labor and newborn infant. Acta Obstet Gynecol Scand 1995; 74: 594–598.
Connor WE, Lowensohn R, Hatcher L: Increased docosahexaenoic acid levels in human newborn infants by administration of sardines and fish oil during pregnancy. Lipids 1996; 31:S183–S187.
van Houwelingen AC, Sorensen JD, Hornstra G, Simonis MM, Boris J, Olsen SF, Secher NJ: Essential fatty acid status in neonates after fish-oil supplementation during late pregnancy. Br J Nutr 1995; 74: 723–731.
Dunstan JA, Mori TA, Barden A, Beilin LJ, Holt PG, Calder PC, Taylor AL, Prescott SL: Effects of n-3 polyunsaturated fatty acid supplementation in pregnancy on maternal and fetal erythrocyte fatty acid composition. Eur J Clin Nutr 2004; 58: 429–437.
Montgomery C, Speake BK, Cameron A, Sattar N, Weaver LT: Maternal docosahexaenoic acid supplementation and fetal accretion. Brit J Nutr 2003; 90: 135–145.
Harris WS, Connor WE, Lindsey S: Will dietary omega-3 fatty acids change the composition of human milk? Am J Clin Nutr 1984; 40: 780–785.
Hawkes JS, Bryan DL, Makrides M, Neumann MA, Gibson RA: A randomized trial of supplementation with docosahexaenoic acid-rich tuna oil and its effects on the human milk cytokines interleukin 1 beta, interleukin 6, and tumor necrosis factor alpha. Am J Clin Nutr 2002; 75: 754–760.
Jensen CL, Maude M, Anderson RE, Heird WC: Effect of docosahexaenoic acid supplementation of lactating women on the fatty acid composition of breast milk lipids and maternal and infant plasma phospholipids. Am J Clin Nutr 2000; 71: 292S-299S.
Urwin HJ, Miles EA, Noakes PS, Kremmyda LS, Vlachava M, Diaper ND, Pérez-Cano FJ, Godfrey KM, Calder PC, Yaqoob P: Salmon consumption during pregnancy alters fatty acid composition and secretory IgA concentration in human breast milk. J Nutr 2012; 142: 1603–1610.
Miles EA, Calder PC: Modulation of immune function by dietary fatty acids. Proc Nutr Soc 1998; 57: 277–292.
Lauritzen L, Brambilla P, Mazzocchi A, Harsløf LB, Ciappolino V, Agostoni C: DHA effects in brain development and function. Nutrients 2016; 8: 6.
Cakiner-Egilmez T: Omega 3 fatty acids and the eye. Insight 2008; 33: 20–25.
Mitchell DC, Litman BJ: Docosahexaenoic acid-containing phospholipids optimally promote rhodopsin activation; in Riemersma RA, Armstrong R, Kelly RW, Wilson R (eds): Essential Fatty Acids and Eicosanoids: Invited Papers from the Fourth International Congress. Champaign, American Oil Chemists’ Society, 1998, pp 154–158.
Zapata-Gonzalez F, Rueda F, Petriz J, Domingo P, Villarroya F, Diaz-Delfin J, de Madariaga MA, Domingo JC: Human dendritic cell activities are modulated by the omega-3 fatty acid, docosahexaenoic acid, mainly through PPAR(gamma):RXR heterodimers: comparison with other polyunsaturated fatty acids. J Leukoc Biol 2008; 84: 1172–1182.
Lefils-Lacourtablaise J, Socorro M, Géloën A, Daira P, Debard C, Loizon E, Guichardant M, Dominguez Z, Vidal H, Lagarde M, Bernoud- Hubac N: The eicosapentaenoic acid metabolite 15-deoxy-δ(12,14)-prostaglandin J3 increases adiponectin secretion by adipocytes partly via a PPARγ-dependent mechanism. PLoS One 2013; 8:e63997.
Woodman RJ, Mori TA, Burke V, Puddey IB, Watts GF, Beilin LJ: Effects of purified eicosapentaenoic and docosahexaenoic acids on glycemic control, blood pressure, and serum lipids in type 2 diabetic patients with treated hypertension. Am J Clin Nutr 2002; 76: 1007– 1015.
Bhaswant M, Poudyal H, Brown L: Mechanisms of enhanced insulin secretion and sensitivity with n-3 unsaturated fatty acids. J Nutr Biochem 2015; 26: 571–584.
Calder PC: Marine omega-3 fatty acids and inflammatory processes: effects, mechanisms and clinical relevance. Biochim Biophys Acta 2015; 1851: 469–484.
Oh DY, Talukdar S, Bae EJ, Imamura T, Morinaga H, Fan W, Li P, Lu WJ, Watkins SM, Olefsky JM: GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010; 142: 687–698.
Bannenberg G, Serhan CN: Specialized proresolving lipid mediators in the inflammatory response: an update. Biochim Biophys Acta 2010; 1801: 1260–1273.
Serhan CN, Yacoubian S, Yang R: Anti-inflammatory and proresolving lipid mediators. Annu Rev Pathol 2008; 3: 279–312.
Mas E, Croft KD, Zahra P, Barden A, Mori TA: Resolvins D1, D2, and other mediators of self-limited resolution of inflammation in human blood following n-3 fatty acid supplementation. Clin Chem 2012; 58: 1476–1484.
Titos E, Clària J: Omega-3-derived mediators counteract obesity-induced adipose tissue inflammation. Prostaglandins Other Lipid Mediat 2013; 107: 77–84.
Weiss GA, Troxler H, Klinke G, Rogler D, Braegger C, Hersberger M: High levels of anti- inflammatory and pro-resolving lipid mediators lipoxins and resolvins and declining docosahexaenoic acid levels in human milk during the first month of lactation. Lipids Health Dis 2013; 12: 89.