Benefits of two Human Milk Oligosaccharides, 2’Fucosyllactose and Lacto-N-Neotetraose in Infant Nutrition

ABSTRACT
Human breast milk contains numerous biomolecules. Human milk oligosaccharides (HMOs) are the third most abundant component of breast milk, after lactose and lipids. Amongst the synthetized HMOs, 2′-fucosyllactose (2′-FL) and lacto-N-neotetraose (LNnT) are widely studied and are considered safe for infant nutrition. Several studies have reported the health benefits of HMOs, which include modulation of the intestinal microbiota, anti-adhesive effect against pathogens, modulation of the intestinal epithelial cell response, and development of the immune system. The amount and diversity of HMOs are determined by the genetic background of the mothers (HMO secretors or non-secretors). The non-secretor mothers secrete lower HMOs than secretor mothers. The breastfed infants of secretor mothers gain more health benefit than those of non-secretor mothers. In conclusion, supplementation of infant formula with 2′-FL and LNnT is a promising innovation for infant nutrition.
INTRODUCTION
According to the recommendations of the World Health Organization, infants must be exclusively breastfed during the first six months of life. Human breast milk provides more than half of the child's nutritional needs during the second year of life [1]. The infants who are formula-fed are more prone to infectious diseases, such as gastroenteritis and acute otitis media, and immune-mediated diseases such as allergy, when compared to the infants who are exclusively breastfed [2]. The composition of breast milk is highly complex, as it contains numerous biomolecules. The human breast milk has 2 types of carbohydrates: lactose and oligosaccharides. Lactose, a major component of human breast milk, has a high nutrition value. Human milk oligosaccharides (HMOs) are the third most abundant component of breast milk, after lactose and lipids. HMOs in the human breast milk are a complex mixture of more than 200 non-digestible and nonnutritional carbohydrates [3]. Among the various compositional differences between human breast milk and cow milk, one of the major differences is the presence of HMOs in human milk, which are virtually absent in cow milk and infant formula [4].
This review discusses the composition of human milk and its biological benefit for infants. Additionally, we also discuss how these beneficial effects can be mimicked if breastfeeding is not possible.
HMOS
The survival advantage of breastfed infants over non-breastfed infants is known since the 1900s. The stool bacterial composition of breastfed infants was reported to be different from that of the formula-fed infants. Additionally, the presence of an unidentified carbohydrate fraction was also reported in human breast milk.
The amount and composition of HMOs vary among women, and also during the lactation period. Generally, the total HMO concentration is higher during the early stages of lactation and decreases within the first three months [5-7]. The HMO content of breast milk after term delivery is higher than that after preterm delivery. The HMO fraction is the third most abundant component in human milk after lactose and lipids, excluding water. The HMO content usually varies between 10–15 grams per liter (g/L) of mature milk (or 1.5–2.3 g/100 kcal, assuming an energy density of human milk of 64 kcal/100 mL) and 20–25 g/L of colostrum [4,8,9]. The HMO content in the human breast milk is more abundant than the protein content, which is typically around 10 g/L or 1.5 g/100 kcal.
The human breast milk contains three major HMO types: fucosylated HMOs (35%–50%), sialylated HMOs (12%–14%), and nonfucosylated neutral HMOs (42%–55%) [10]. Fucosylated HMOs include 2′-fucosyllactose (2′-FL), while non-fucosylated neutral HMOs include lacto-N-neotetraose (LNnT). The neutral HMOs account for more than 75% of the total HMOs in the human breast milk. The most abundant HMO is the 2′-FL, which constitutes nearly 30% of the total HMOs among secretor mothers. The factors that account for the variability in the secretion of HMOs include the geographical origin and genetic background of the mothers. The HMO content in the breast milk varies among women. Furthermore, the amount and diversity of HMOs are determined by the genetic background of the mothers (Table 1). The Lewis antigen system is a human blood group system based upon the genes on chromosome 19 p13.3 (fucosyltransferase [FUT] 3 or Lewis gene) and 19q13.3, (FUT2 or secretor gene). Both genes are expressed in the glandular epithelia. The dominant allele of these genes encodes enzymes with FUT activity, while the recessive alleles are non-functional. Globally, it is estimated that about 20% of mothers secrete lower amount of HMOs (non-secretor mothers) than the secretor mothers. An earlier study evaluated the HMO content during the lactation period based on the concentration of 2′-FL. The study suggested that the concentration of LNnT and lacto-N-tetraose (LNT) is co-regulated by the FUT2-dependent 2′-FL concentration. The concentration of 2′-FL was positively correlated with the concentration of LNnT and was negatively correlated with the concentration of LNT [11].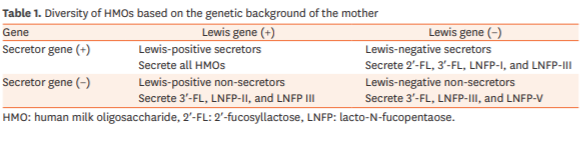
One of the factors that affect the composition of primary gut microbiome in infants is feeding. The intestine of a neonate is already colonized at birth. A healthy gut microbiome protects the host against pathogens by various mechanisms such as enhancing the immune development and stimulating the digestive and metabolic functions. Other factors that affect the colonization of the gut include the gestation period, mode of delivery, environment, and medication. Dysbiosis during early life is a risk factor for immune-mediated diseases, such as allergy and asthma, and intestinal and metabolic diseases.
The gut microbiome of infants born through cesarean section (c-section) is different from that of infants born through vaginal birth. The number of bifidobacteria in the intestinal microbiome of infants born through c-section may take up to 6 months to reach the number of bifidobacteria observed in the intestinal microbiome of infants born through vaginal delivery [12]. The infants born through c-section have a higher risk to develop allergic disease (odd ratio [OR], 1.23l; p=0.007), obesity (58% higher) and type 1 diabetes mellitus (adjusted OR, 1.19; 95% confidence interval [CI], 1.04–1.36; p=0.01) compared to the infants born through vaginal birth [13-15]. Additionally, infant feeding can influence the microbiome composition. The difference in the microbiome composition between the breastfed infants and formula-fed infants is mainly due to the absence of HMOs in cow milk. The consumption of infant formula not supplemented with HMOs will result in a microbiome composition that is poor in bifidobacteria, which can affect the immune development [16]. Additionally, the exposure to antibiotics during the first year of life can also affect the gut microbiome composition. The administration of antibiotics earlier in life and the frequency of antibiotic administration affect the gut microbiome composition [17]. Infants treated with antibiotics are at a higher risk to develop cow's milk protein allergy (OR, 1.26; 95% CI, 1.20–1.33) [18], obesity (32.4 vs. 18.2%, p=0.002) [19], asthma (OR, 7.77; 95% CI, 6.25–9.65) [20], otitis (OR, 1.30; 95% CI, 1.040–1.63) [21], inflammatory bowel disease (OR, 5.51; 95% CI, 1.66–18.2) [22], and diabetes (OR, 1.08; 95% CI, 1.05–1.11) than infants not treated with antibiotics [23]. Antibiotics kill not only the pathogen but also the beneficial bacteria, which compromises immune homeostasis and deregulates metabolism. Additionally, antibiotic administration is also associated with the development of antibiotic resistance [24]. Exposure to proton pump inhibitors (PPIs) and H2 receptor antagonists can also increase the risk to develop immunemediated diseases, such as atopic dermatitis, asthma, and allergic rhinitis [25].
The establishment of a bifidobacteria-predominant microbiota in the infants who are exclusively breastfed by non-secretor mothers is slower when compared to that in infants who are breastfed by secretor mothers [26]. The infants who are breastfed by secretor mothers are less likely to develop diarrhea, specifically Campylobacter gastroenteritis, when compared to those who are breastfed by non-secretor mothers [27]. Several studies have suggested that the health outcome of infants who are exclusively breastfed by secretor mothers is better than that of infants who are breastfed by non-secretor mothers.
The HMO composition also represents the blood group characteristics. The presence of different neutral oligosaccharides in the human breast milk depends on the activity of specific enzymes (FUTs), which are related to the Lewis blood group [8,28].
HEALTH BENEFIT OF HUMAN MILK OLIGOSACCHARIDES
Several studies have reported the beneficial effects of HMOs that include modification of the intestinal microbiota, anti-adhesive effect against pathogens, modulation of the intestinal epithelial cell response, and development of the immune system. We will discuss each of these effects further.
Modulation of intestinal microbiota
HMOs are intrinsic components that affect the gut microbiota by providing an energy source for the beneficial intestinal bacteria. Additionally, HMOs affect the health of the host by serving as a decoy receptor for the opportunistic pathogens in the mucosal surface [29]. One study reported that none of the selected Enterobacteriaceae strains exhibited growth on a medium containing 2′-FL, 6′-sialyllactose or LNnT as a carbohydrate source. However, several strains were capable of utilizing galacto-oligosaccharides (GOS), maltodextrin, and monosaccharide and disaccharide components of HMOs for their growth [30]. The enriched fecal consortia also did not exhibit growth on a medium containing 2′-FL or 6′-sialyllactose, but exhibited limited growth on a medium containing LNnT [30]. Several in vitro studies have demonstrated that HMOs promote the growth of certain but not all Bifidobacterium [31]. Bifidobacterium longum subsp. Bifidobacterium infantis exhibit good growth on medium supplemented with HMOs, including 2′-FL, as the sole source of carbohydrate [32-35]. Over time, B. infantis consumes all HMOs including its monosaccharide and disaccharide metabolites [34].
The growth of Bifidobacterium bifidum is slower than that of B. infantis in the presence of HMOs. Additionally, certain B. longum strains metabolize fucosylated HMOs [4,34,36]. The Bifidobacterium kashiwanohense strain exhibits growth in the presence of 2′-FL and 3′-FL [35]. HMOs are a preferred substrate for B. infantis. Other bifidobacteria may reduce the nutrients available for potentially harmful bacteria and limit their growth. Additionally, B. infantis produces short-chain fatty acids (SCFAs), which favor the growth of commensal bacteria and not pathogenic bacteria [37]. A study reported that among the 24 probiotic strains, only B. longum subsp. B. infantis ATCC 15697 and B. infantis M-63 were able to ferment 3′-sialyllactose, 6′-sialyllactose, 2′-FL, and 3′-FL [38].
When infants are fed with a formula supplemented with 2′-FL and LNnT, they develop a distinctive stool bacterial profile that is more similar to that of the breastfed infants compared to the infants that are fed with a formula not supplemented with prebiotics. The bacterial diversity of infants at the age of 3 months exhibited increased colonization with beneficial bifidobacteria and decreased colonization with pathogenic bacteria [39].
Antiadhesive properties
HMOs improve the host defense mechanism by strengthening the gut barrier function [40]. The HMO, 2′-FL inhibits Campylobacter jejuni infection and C. jejuni-associated mucosal inflammation [41]. An in vitro study demonstrated that 2′-FL attenuates C. jejuni invasion by 80% and inhibits the release of mucosal pro-inflammatory signals. A study on mouse model revealed that the ingestion of 2′-FL inhibits the C. jejuni colonization by 80%, weight loss by 5%, intestinal inflammation, and induction of inflammatory signaling molecules [42]. A prospective study on infants suggested that the beneficial effect of 2′-FL includes a reduction in the number of episodes of C. jejuni-associated diarrhea [43]. LNnT was reported to reduce the abundance of Streptococcus pneumoniae in the lungs of an animal model [44]. HMOs may function as a decoy receptor for group B Streptococcus [45].
HMOs reduce preterm mortality and morbidity by modulating the gut microbiome to protect against necrotizing enterocolitis, candidiasis, and several immune-related diseases [46]. LNnT reduces the risk of developing necrotizing enterocolitis in preterm infants [47]. Similarly, 2′-FL has also been reported to exhibit beneficial effect against necrotizing enterocolitis [48].
Modulators of intestinal cell response
HMOs are able to directly affect the intestinal cell response by reducing the cell growth and by inducing differentiation and apoptosis [49]. Intestinal health and barrier function are considered to be the first line of defense in innate immunity [50]. HMOs have been reported to increase the intestinal cell maturation [50].
Immune modulators
One of the important properties of HMOs is the immunomodulation. HMOs directly modulate the gene expression of intestinal cells, leading to changes in the expression of cell surface glycans and other cell responses [51]. HMOs modulate lymphocyte cytokine production and enable a more balanced TH1/TH2 response. An increasing number of in vitro studies suggest that HMOs exert microbiota-independent effects by directly modulating the immune response and by regulating the immune cell population and cytokine secretion [52]. HMOs may either act locally on the mucosa-associated lymphoid tissue or act at a systemic level [4]. The plasma concentration of inflammatory cytokines in the breastfed infants and infants fed with experimental formula supplemented with 2′-FL was markedly lower than that in the infants fed with control formula supplemented with galacto-oligosaccharides [53]. These data indicate that infants fed with a formula supplemented with 2′-FL exhibit lower plasma inflammatory cytokine profiles, which is similar to those of a breastfed reference group [53]. HMOs were more effective than non-human prebiotic oligosaccharides in modulating the systemic and gastrointestinal immune cell responses in pigs [54]. These altered immune cell populations may mediate the rotavirus infection susceptibility [54]. The symptoms of food allergy are reduced by 2′-FL through induction of interleukin-10+ T-regulatory cells and through indirect stabilization of mast cells [55].
HMOs, especially 2′-FL, directly inhibit the lipopolysaccharide-mediated inflammation during enterotoxigenic Escherichia coli invasion of T84 and H4 intestinal epithelial cells through attenuation of CD14 induction [56]. CD14 expression mediates the lipopolysaccharide-Toll-like receptor 4 stimulation of a part of the macrophage migration inhibitory factors inflammatory pathway by suppressing the cytokine signaling 2/signal transducer and by activating the transcription factor 3/nuclear factor-κB. The direct inhibition of inflammation supports the role of HMOs as a stimulator of the innate immune system [56]. Two-year-old children who were born through c-section and fed on an infant formula supplemented with 2′-FL had a lower risk of developing immunoglobulin E-associated allergies compared to those fed unsupplemented formula [11].
BRAIN DEVELOPMENT
HMOs and their metabolic products, such as sialic acid, have a role in brain development, neuronal transmission, and synaptogenesis. HMOs are a source of sialic acid, which is an essential nutrient for optimal brain development and cognition [57,58]. L-fucose and 2′-FL stimulate brain development of [59]. Dietary 2′-FL affects cognitive domains and improves learning and memory in rodents [60]. The HMOs 3′-sialyllactose and 6′-sialyllactose support normal microbial communities and behavioral responses during stress by modulating the gut-brain axis [61].
SAFETY AND CLINICAL OUTCOMES
Oligosaccharides were identified as the bifidogenic factor in human milk in the 1930s. The most abundant oligosaccharides in the human breast milk were discovered and characterized in 1954. However, the industrial production of some of the HMOs was recently achieved. The molecular structure of industrially produced 2′-FL and LNnT is identical to that of the oligosaccharides present in the human breast milk. Unlike probiotics, HMOs are resistant to pasteurization and freeze-drying [62,63].
Two intervention studies tested the safety of an infant formula supplemented with approximately 0.2 g/L 2′-FL [64,65]. When combined with GOS, 2′-FL (0.2 or 1.0 g/L) did not affect the growth parameters, was well tolerated, and did not influence the stool frequency or consistency [64]. There was no difference in the frequency of adverse events between the infants who were fed with a formula supplemented with 2′-FL and those fed with a control infant formula supplemented with GOS. Similarly, when infants were fed with an infant formula supplemented with fructo-oligosaccharides (FOS) and 2′-FL (0.2 g/L) for a period of approximately 1 month, the formula was well tolerated [65].
Janas et al. [66] fed the infants with a formula supplemented with 2′-FL or LNnT from less than 14 days of life to the age of 6 months (4 months exclusively). The formula was reported to be safe and well-tolerated, and supported an age-appropriate growth [66]. Additionally, there was no difference in the stool consistency and stool frequency between the 2′-FL-fed group and the LNnT-fed group, except at 2 months when stools were softer in the 2′-FL-fed group [66]. In infants aged less than 8 days of life at inclusion, Kajzer et al. [65] showed excellent tolerance fed of a formula supplemented with 2′-FL and FOS for 1 month. Prieto [67] demonstrated that administration of LNnT to older infants and young children, aged 6–24 months, for a period of 4 months resulted in normal growth. This was the only study that used a follow-up formula.
Puccio et al. [39] analyzed the incidence of different health outcomes as secondary outcomes, and observed a reduced incidence of bronchitis at 4, 6 and 12 months, reduced incidence of lower respiratory tract infections through 12 months, reduced use of antipyretics through 4 months, reduced use of antibiotics through 6 and 12 months with protective effects that continued after 6 months post-intervention period of an infant formual supplemented with 2′-FL and LNnT.
The US Food and Drug Administration categorized three HMOs as generally regarded as safe (GRAS notice n° 650): 2′-O-fucosyllactose, 2'-FL, and LNnT. In the European Union, HMOs are considered as novel foods and the oligosaccharides, 2'-FL and LNnT, and their combination passed the safety assessment [29]. The European Food Safety Authority (EFSA) positively assessed 2′-FL on June 29, 2015, based on the scientific and technical information and reported that [68]: “2′-FL is safe for infants (up to one year of age) when added to infant and follow-on formulae, in combination with LNnT, at concentrations up to 1.2 g/L of 2′-FL and up to 0.6 g/L of LNnT, at a ratio of 2:1 in the reconstituted formulae. 2′-FL is safe for young children (older than 1 year of age) when added to follow-on and young-child formulae, at concentrations up to 1.2 g/L of 2′-FL (alone or in combination with LNnT, at concentrations up to 0.6 g/L, at a ratio of 2:1).” The HMO, 2′-FL is reported to contribute to the infant's and child's health. Amongst the HMOs, 2′-FL and LNnT are widely studied and have a chemically simple structure. These two HMOs are more abundant in the human breast milk compared to other HMOs. Additionally, these 2 HMOs can be produced on an industrial scale. Hence, these HMOs can be used as supplements to infant formula.
CONCLUSION
HMOs can serve as soluble decoy receptors that block the attachment of viral, bacterial, or protozoan parasitic pathogens to the epithelial cell surface receptors, which may aid in preventing infectious diseases. HMOs are also antimicrobials that act as bacteriostatic or bactericidal agents. Additionally, HMOs enhance host epithelial and immune cell responses in the neonate.
Although the functions of HMOs were known previously, a strategy for industrial production was not available. Hence, non-human milk oligosaccharides, mainly FOS and GOS, were used as an alternative supplement for the infant formula. Currently, 2′-FL is added to the infant formula as the industrial production capacity has increased. The industrial production of other HMOs, such as LNnT is still limited. Hence, LNnT is not routinely used as a supplement to the infant formula.
The gastrointestinal microbiome of infants fed with a formula supplemented with 2′-FL is similar to that of the infants who are exclusively breastfed. There have been no adverse effects reported till date for 2′-FL. Clinical studies have demonstrated that infants fed on a formula supplemented with 2′-FL exhibit a normal growth pattern and normal defecation. Therefore, it can be concluded that 2′-FL is a safe supplementation for infant formula.
REFERENCES
1. World Health Organization. Maternal, newborn, child and adolescent health [Internet]. Geneva: World Health Organization; [cited 2018 Oct 31]. Available from: http://www.who.int/maternal_child_adolescent/ topics/child/nutrition/breastfeeding/en/.
2. ESPGHAN Committee on Nutrition, Agostoni C, Braegger C, Decsi T, Kolacek S, Koletzko B, et al. Breast-feeding: a commentary by the ESPGHAN Committee on Nutrition. J Pediatr Gastroenterol Nutr 2009;49:112-25. PUBMED | CROSSREF
3. Lönnerdal B, Hernell O. An opinion on “staging” of infant formula- a developmental perspective on infant feeding. J Pediatr Gastroenterol Nutr 2016;62:9-21. PUBMED | CROSSREF
4. Bode L. Human milk oligosaccharides: every baby needs a sugar mama. Glycobiology 2012;22:1147-62. PUBMED | CROSSREF
5. Xu G, Davis JC, Goonatilleke E, Smilowitz JT, German JB, Lebrilla CB. Absolute quantitation of human milk oligosaccharides reveals phenotypic variations during lactation. J Nutr 2017;147:117-24. PUBMED | CROSSREF
6. Chaturvedi P, Warren CD, Altaye M, Morrow AL, Ruiz-Palacios G, Pickering LK, et al. Fucosylated human milk oligosaccharides vary between individuals and over the course of lactation. Glycobiology 2001;11:365-72. PUBMED | CROSSREF
7. Thurl S, Munzert M, Henker J, Boehm G, Müller-Werner B, Jelinek J, et al. Variation of human milk oligosaccharides in relation to milk groups and lactational periods. Br J Nutr 2010;104:1261-71. PUBMED | CROSSREF
8. Kunz C, Kuntz S, Rudloff S. Bioactivity of human milk oligosaccharides. In: Moreno FM, Sanz ML, eds. Food Oligosaccharides: Production, Analysis and Bioactivity. 1st ed. Chichester: John Wiley & Sons, Ltd; 2014: 5-20. CROSSREF
9. Zivkovic AM, German JB, Lebrilla CB, Mills DA. Human milk glycobiome and its impact on the infant gastrointestinal microbiota. Proc Natl Acad Sci U S A 2011;108 Suppl 1:4653-8. PUBMED | CROSSREF
10. Smilowitz JT, Lebrilla CB, Mills DA, German JB, Freeman SL. Breast milk oligosaccharides: structurefunction relationships in the neonate. Annu Rev Nutr 2014;34:143-69. PUBMED | CROSSREF
11. Sprenger N, Lee LY, De Castro CA, Steenhout P, Thakkar SK. Longitudinal change of selected human milk oligosaccharides and association to infants' growth, an observatory, single center, longitudinal cohort study. PLoS One 2017;12:e0171814. PUBMED | CROSSREF
12. Grönlund MM, Lehtonen OP, Eerola E, Kero P. Fecal microflora in healthy infants born by different methods of delivery: permanent changes in intestinal flora after cesarean delivery. J Pediatr Gastroenterol Nutr 1999;28:19-25. PUBMED | CROSSREF
13. Renz-Polster H, David MR, Buist AS, Vollmer WM, O'Connor EA, Frazier EA, et al. Caesarean section delivery and the risk of allergic disorders in childhood. Clin Exp Allergy 2005;35:1466-72. PUBMED | CROSSREF
14. Goldani HA, Bettiol H, Barbieri MA, Silva AA, Agranonik M, Morais MB, et al. Cesarean delivery is associated with an increased risk of obesity in adulthood in a Brazilian birth cohort study. Am J Clin Nutr 2011;93:1344-7. PUBMED | CROSSREF
15. Cardwell CR, Stene LC, Joner G, Cinek O, Svensson J, Goldacre MJ, et al. Caesarean section is associated with an increased risk of childhood-onset type 1 diabetes mellitus: a meta-analysis of observational studies. Diabetologia 2008;51:726-35. PUBMED | CROSSREF
16. Harmsen HJ, Wildeboer-Veloo AC, Raangs GC, Wagendorp AA, Klijn N, Bindels JG, et al. Analysis of intestinal flora development in breast-fed and formula-fed infants by using molecular identification and detection methods. J Pediatr Gastroenterol Nutr 2000;30:61-7. PUBMED | CROSSREF
17. Yasmin F, Tun HM, Konya TB, Guttman DS, Chari RS, Field CJ, et al. Caesarean section, formula feeding, and infant antibiotic exposure: separate and combined impacts on gut microbial changes in later infancy. Front Pediatr 2017;5:200. PUBMED | CROSSREF
18. Mikkelsen KH, Knop FK, Vilsbøll T, Frost M, Hallas J, Pottegård A. Use of antibiotics in childhood and risk of type 1 diabetes: a population-based case-control study. Diabet Med 2017;34:272-7. PUBMED | CROSSREF
19. Azad MB, Bridgman SL, Becker AB, Kozyrskyj AL. Infant antibiotic exposure and the development of childhood overweight and central adiposity. Int J Obes 2014;38:1290-8. PUBMED | CROSSREF
20. Wu P, Feldman AS, Rosas-Salazar C, James K, Escobar G, Gebretsadik T, et al. Relative importance and additive effects of maternal and infant risk factors on childhood asthma. PLoS One 2016;11:e0151705. PUBMED | CROSSREF
21. Ilić K, Jakovljević E, Skodrić-Trifunović V. Social-economic factors and irrational antibiotic use as reasons for antibiotic resistance of bacteria causing common childhood infections in primary healthcare. Eur J Pediatr 2012;171:767-77.
22. Hviid A, Svanström H, Frisch M. Antibiotic use and inflammatory bowel diseases in childhood. Gut 2011;60:49-54. PUBMED | CROSSREF
23. Kronman MP, Zaoutis TE, Haynes K, Feng R, Coffin SE. Antibiotic exposure and IBD development among children: a population-based cohort study. Pediatrics 2012;130:e794-803. PUBMED | CROSSREF
24. Francino MP. Antibiotics and human gut microbiome: dysbioses and accumulation of resistances. Front Microbiol 2016;6:1543. PUBMED | CROSSREF
25. Mulder B, Schuiling-Veninga CC, Bos HJ, De Vries TW, Jick SS, Hak E. Prenatal exposure to acidsuppressive drugs and the risk of allergic diseases in the offspring: a cohort study. Clin Exp Allergy 2014;44:261-9. PUBMED | CROSSREF
26. Lewis ZT, Totten SM, Smilowitz JT, Popovic M, Parker E, Lemay DG, et al. Maternal fucosyltransferase 2 status affects the gut bifidobacterial communities of breastfed infants. Microbiome 2015;3:13. PUBMED | CROSSREF
27. Morrow AL, Ruiz-Palacios GM, Altaye M, Jiang X, Guerrero ML, Meinzen-Derr JK, et al. Human milk oligosaccharides are associated with protection against diarrhea in breast-fed infants. J Pediatr 2004;145:297-303. PUBMED | CROSSREF
28. Bode L. The functional biology of human milk oligosaccharides. Early Hum Dev 2015;91:619-22. PUBMED | CROSSREF
29. Salminen S. Regulatory aspect of human milk oligosaccharides. Nestle Nutr Inst Workshop Ser 2017;88:161-70. PUBMED | CROSSREF
30. Hoeflinger JL, Davis SR, Chow J, Miller MJ. In vitro impact of human milk oligosaccharides on Enterobacteriaceae growth. J Agric Food Chem 2015;63:3295-302. PUBMED | CROSSREF
31. Bode L, Kuhn L, Kim HY, Hsiao L, Nissan C, Sinkala M, et al. Human milk oligosaccharide concentration and risk of postnatal transmission of HIV through breastfeeding. Am J Clin Nutr 2012;96:831-9. PUBMED | CROSSREF
32. LoCascio RG, Ninonuevo MR, Freeman SL, Sela DA, Grimm R, Lebrilla CB, et al. Glycoprofiling of bifidobacterial consumption of human milk oligosaccharides demonstrates strain specific, preferential consumption of small chain glycans secreted in early human lactation. J Agric Food Chem 2007;55:8914-9. PUBMED | CROSSREF
33. Marcobal A, Barboza M, Froehlich JW, Block DE, German JB, Lebrilla CB, et al. Consumption of human milk oligosaccharides by gut-related microbes. J Agric Food Chem 2010;58:5334-40. PUBMED | CROSSREF
34. Asakuma S, Hatakeyama E, Urashima T, Yoshida E, Katayama T, Yamamoto K, et al. Physiology of consumption of human milk oligosaccharides by infant gut-associated bifidobacteria. J Biol Chem 2011;286:34583-92. PUBMED | CROSSREF
35. Bunesova V, Lacroix C, Schwab C. Fucosyllactose and L-fucose utilization of infant Bifidobacterium longum and Bifidobacterium kashiwanohense. BMC Microbiol 2016;16:248. PUBMED | CROSSREF
36. Garrido D, Ruiz-Moyano S, Kirmiz N, Davis JC, Totten SM, Lemay DG, et al. A novel gene cluster allows preferential utilization of fucosylated milk oligosaccharides in Bifidobacterium longum subsp. longum SC596. Sci Rep 2016;6:35045. PUBMED | CROSSREF
37. Gibson GR, Wang X. Regulatory effects of bifidobacteria on the growth of other colonic bacteria. J Appl Bacteriol 1994;77:412-20. PUBMED | CROSSREF
38. Thongaram T, Hoeflinger JL, Chow J, Miller MJ. Human milk oligosaccharide consumption by probiotic and human-associated bifidobacteria and lactobacilli. J Dairy Sci 2017;100:7825-33. PUBMED | CROSSREF
39. Puccio G, Alliet P, Cajozzo C, Janssens E, Corsello G, Sprenger N, et al. Effects of infant formula with human milk oligosaccharides on growth and morbidity: a randomized multicenter trial. J Pediatr Gastroenterol Nutr 2017;64:624-31.
40. Angeloni S, Ridet JL, Kusy N, Gao H, Crevoisier F, Guinchard S, et al. Glycoprofiling with micro-arrays of glycoconjugates and lectins. Glycobiology 2005;15:31-41. PUBMED | CROSSREF
41. Yu ZT, Nanthakumar NN, Newburg DS. The human milk oligosaccharide 2′-fucosyllactose quenches Campylobacter jejuni-induced inflammation in human epithelial cells HEp-2 and HT-29 and in mouse intestinal mucosa. J Nutr 2016;146:1980-90. PUBMED | CROSSREF
42. Ruiz-Palacios GM, Cervantes LE, Ramos P, Chavez-Munguia B, Newburg DS. Campylobacter jejuni binds intestinal H(O) antigen (Fucα1, 2Galβ1, 4GlcNAc), and fucosyloligosaccharides of human milk inhibit its binding and infection. J Biol Chem 2003;278:14112-20. PUBMED | CROSSREF
43. Morrow AL, Ruiz-Palacios GM, Jiang X, Newburg DS. Human-milk glycans that inhibit pathogen binding protect breast-feeding infants against infectious diarrhea. J Nutr 2005;135:1304-7. PUBMED | CROSSREF
44. Idänpään-Heikkilä I, Simon PM, Zopf D, Vullo T, Cahill P, Sokol K, et al. Oligosaccharides interfere with the establishment and progression of experimental pneumococcal pneumonia. J Infect Dis 1997;176:704-12. PUBMED | CROSSREF
45. Lin AE, Autran CA, Szyszka A, Escajadillo T, Huang M, Godula K, et al. Human milk oligosaccharides inhibit growth of group B Streptococcus. J Biol Chem 2017;292:11243-9. PUBMED | CROSSREF
46. Moukarzel S, Bode L. Human milk oligosaccharides and the preterm infant: a journey in sickness and in health. Clin Perinatol 2017;44:193-207. PUBMED | CROSSREF
47. Autran CA, Kellman BP, Kim JH, Asztalos E, Blood AB, Spence EC, et al. Human milk oligosaccharide composition predicts risk of necrotising enterocolitis in preterm infants. Gut 2018;67:1064-70. PUBMED | CROSSREF
48. Good M, Sodhi CP, Yamaguchi Y, Jia H, Lu P, Fulton WB, et al. The human milk oligosaccharide 2′-fucosyllactose attenuates the severity of experimental necrotising enterocolitis by enhancing mesenteric perfusion in the neonatal intestine. Br J Nutr 2016;116:1175-87. PUBMED | CROSSREF
49. Kuntz S, Kunz C, Rudloff S. Oligosaccharides from human milk induce growth arrest via G2/M by influencing growth-related cell cycle genes in intestinal epithelial cells. Br J Nutr 2009;101:1306-15. PUBMED | CROSSREF
50. Holscher HD, Davis SR, Tappenden KA. Human milk oligosaccharides influence maturation of human intestinal Caco-2Bbe and HT-29 cell lines. J Nutr 2014;144:586-91. PUBMED | CROSSREF
51. Kulinich A, Liu L. Human milk oligosaccharides: The role in the fine-tuning of innate immune responses. Carbohydr Res 2016;432:62-70. PUBMED | CROSSREF
52. Donovan SM, Comstock SS. Human milk oligosaccharides influence neonatal mucosal and systemic immunity. Ann Nutr Metab 2016;69 Suppl 2:42-51. PUBMED | CROSSREF
53. Goehring KC, Marriage BJ, Oliver JS, Wilder JA, Barrett EG, Buck RH. Similar to those who are breastfed, infants fed a formula containing 2′-fucosyllactose have lower inflammatory cytokines in a randomized controlled trial. J Nutr 2016;146:2559-66. PUBMED | CROSSREF
54. Comstock SS, Li M, Wang M, Monaco MH, Kuhlenschmidt TB, Kuhlenschmidt MS, et al. Dietary human milk oligosaccharides but not prebiotic oligosaccharides increase circulating natural killer cell and mesenteric lymph node memory T cell populations in noninfected and rotavirus-infected neonatal piglets. J Nutr 2017;147:1041-7. PUBMED | CROSSREF
55. Castillo-Courtade L, Han S, Lee S, Mian FM, Buck R, Forsythe P. Attenuation of food allergy symptoms following treatment with human milk oligosaccharides in a mouse model. Allergy 2015;70:1091-102. PUBMED | CROSSREF
56. He Y, Liu S, Kling DE, Leone S, Lawlor NT, Huang Y, et al. The human milk oligosaccharide 2′-fucosyllactose modulates CD14 expression in human enterocytes, thereby attenuating LPS-induced inflammation. Gut 2016;65:33-46.
57. Bienenstock J, Buck RH, Linke H, Forsythe P, Stanisz AM, Kunze WA. Fucosylated but not sialylated milk oligosaccharides diminish colon motor contractions. PLoS One 2013;8:e76236. PUBMED | CROSSREF
58. Jacobi SK, Yatsunenko T, Li D, Dasgupta S, Yu RK, Berg BM, et al. Dietary isomers of sialyllactose increase ganglioside sialic acid concentrations in the corpus callosum and cerebellum and modulate the colonic microbiota of formula-fed piglets. J Nutr 2016;146:200-8. PUBMED | CROSSREF
59. Matthies H, Staak S, Krug M. Fucose and fucosyllactose enhance in-vitro hippocampal long-term potentiation. Brain Res 1996;725:276-80. PUBMED | CROSSREF
60. Vázquez E, Barranco A, Ramírez M, Gruart A, Delgado-García JM, Martínez-Lara E, et al. Effects of a human milk oligosaccharide, 2′-fucosyllactose, on hippocampal long-term potentiation and learning capabilities in rodents. J Nutr Biochem 2015;26:455-65. PUBMED | CROSSREF
61. Tarr AJ, Galley JD, Fisher SE, Chichlowski M, Berg BM, Bailey MT. The prebiotics 3′Sialyllactose and 6′Sialyllactose diminish stressor-induced anxiety-like behavior and colonic microbiota alterations: evidence for effects on the gut-brain axis. Brain Behav Immun 2015;50:166-77. PUBMED | CROSSREF
62. Hahn WH, Kim J, Song S, Park S, Kang NM. The human milk oligosaccharides are not affected by pasteurization and freeze-drying. J Matern Fetal Neonatal Med 2019;32:985-91. PUBMED | CROSSREF
63. Daniels B, Coutsoudis A, Autran C, Amundson Mansen K, Israel-Ballard K, Bode L. The effect of simulated flash heating pasteurisation and Holder pasteurisation on human milk oligosaccharides. Paediatr Int Child Health 2017;37:204-9. PUBMED | CROSSREF 64. Marriage BJ, Buck RH, Goehring KC, Oliver JS, Williams JA. Infants fed a lower calorie formula with 2′FL show growth and 2′FL uptake like breast-fed infants. J Pediatr Gastroenterol Nutr 2015;61:649-58. PUBMED | CROSSREF
65. Kajzer J, Oliver J, Marriage B. Gastrointestinal tolerance of formula supplemented with oligosaccharides. FASEB J 2016;30 Suppl:671.4.
66. Janas B, Wernimont S, Gosoniu L, Northington R. Clinical safety of a new starter infant formula containing 2 human milk oligosaccharides (HMOs), complete report. Nestlé Nutrition R&D Clinical Study Summary; November 2015. Vevey, Switzerland: Nestlé; 2015.
67. Prieto PA. In vitro and clinical experiences with a human milk oligosaccharide, lacto-N-neoTetraose, and fructooligosaccharides. Food Foods Ingredients J Jpn 2005;210:1018-30.
68. EFSA Panel on Dietetic Products, Nutrition, and Allergies (NDA). Safety of 2′-O-fucosyllactose as a novel food ingredient pursuant to regulation (EC) No 258/97. EFSA J 2015;13:4184.
If you liked this post you may also like



