Advancing the Management of Cow’s Milk Protein Allergy with Human Milk Oligosaccharides: Priming the Immune System
Meeting Summary
Yvan Vandenplas summarised clinical trials of HMOs in CMPA to date, highlighting data on growth, tolerance, and reduction in infection risk achieved with HMO-supplemented specialty formula. The major role that HMOs play in shaping the gut microbiome in early infancy was discussed by Heine. He presented recent data showing how supplementation of standard or extensively hydrolysed infant formula with two HMOs, 2’-fucosyllactose (2’FL) and lacto-N-neotetraose (LNnT), can shift the gut microbiome in CMPA closer to the profile of breastfed infants. Nutten outlined how 2’FL and LNnT can beneficially modulate Type 2 immune responses, which may have important implications for both allergy prevention and treatment strategies.
The Clinical Relevance of Human Milk Oligosaccharides in Infants with Cow’s Milk Protein Allergy
Yvan Vandenplas
Risk factors for CMPA in infants include genetic background, mode of delivery, feeding, and medication, all of which are intrinsically linked to the gut microbiome. Two components of breast milk play a major role in delivering immune support: HMOs, which are virtually absent in cow’s milk and unsupplemented infant formula milk, and lactose.1-4 Lactose acts by upregulating the immune defences, while HMOs act as a major substrate for Bifidobacterium, enhancing levels of short-chain fatty acids (SCFAs), in particular, butyrate.4-7
Lactose and HMOs therefore work synergistically to stimulate development of a well-balanced gut microbiome in breast-fed infants, explained Vandenplas. Of the more than 200 different HMOs identified so far in human breast milk, Vandenplas described non-fucosylated HMOs (e.g., LNnT) and fucosylated HMOs (e.g., 2’FL) as the two key categories.8-10 HMOs confer a protective effect in allergy via modulation of mucosal immunity. In a study of infants born by caesarean section, those fed breast milk containing higher levels of 2’FL had a lower incidence of IgE-associated atopic eczema compared to those receiving milk from ‘non-secretor’ mothers with lower 2’FL levels (14% versus 43%; p=0.095).11
Observational studies have revealed differences in the composition of the gut microbiota between allergic and non-allergic infants, which are present before any manifestation of disease. This clearly shows that dysbiosis is primary to CMPA and not secondary, stressed Vandenplas.12,13 Breast milk contains HMOs, which drive development of a healthy gut microbiota. Alterations during this critical development period increase the risk of the infant developing allergic disease. Infants with CMPA also show a higher propensity for developing infections.14
Currently, the recommended first-choice option in the management of CMPA is an extensively hydrolysed formula (EHF).15 An amino-acid based formula (AAF) may also be indicated in certain cases.16 Vandenplas reviewed clinical data that support a key role for HMOs added to these specialty formulas in the treatment of CMPA. Studies have shown that whey-based extensively hydrolysed formulas (w-EHF) supplemented with HMOs (Althéra® HMO, Nestlé, Vevey, Switzerland) are well tolerated and support normal growth in infants with CMPA. The hypoallergenicity of w-EHF containing additional 2’FL and LNnT has also been confirmed in infants and children with CMPA, meeting guidelines laid down by the American Academy of Pediatrics (AAP).17,18 The test formula supplemented with HMOs was well tolerated and levels of 2’FL and LNnT in w-EHF proved comparable to those found in breast milk.17,19,20 In the CINNAMON European study of nearly 200 infants with CMPA, confirmed growth was also achieved in infants with CMPA being fed w-EHF with added 2’FL and LNnT.21-23
HMO-supplemented formula may confer additional clinical benefits in infants with CMPA and Vandenplas highlighted further recent evidence from the CINNAMON study. Secondary analysis of the CINNAMON study revealed a reduction in the risk of infections, including respiratory tract infections and otitis media, in infants who received HMO-supplemented formula (n=94) versus control (n=96).21-23 Infants with CMPA fed w-EHF formula with additional 2’FL and LNnT showed a clinically significant reduction of 23% in lower respiratory tract infection episodes per month, as well as a statistically significant reduction of 42% in monthly upper respiratory tract infections.21-22 For upper respiratory tract infections in particular, w-EHF with 2’FL and LNnT was associated with a clear reduction in the rate of respiratory infections per month of study formula use, with a hazard ratio (HR) of 0.58 (p=0.003).21-22 Similar results have been seen in healthy infants fed with 2’FL and LNnT supplemented formula, with a significant relative risk (RR) reduction of 44% for reported lower respiratory tract infections.23 Collectively, these data show that the addition of HMOs to w-EHF results in a clinically significant reduction in the risk of respiratory tract infections, Vandenplas concluded.
This reduction in infection risk also led to a clinically significant trend towards decreased use of antibiotic (RR: -20.3%) and antipyretic (RR: -24.9%) medications in infants with CMPA fed w-EHF supplemented with 2’FL and LNnT.21-22 Lower rates of infection-related medication use with HMOs may be linked to gut microbiota community types, suggested Vandenplas. Notably, healthy infants with a high bifidobacteria-dominated gut at 3 months have been shown to be less likely to require antibiotics during the first year of life.24 These clinical benefits may be extended to formula-fed infants by using HMO supplementation to achieve a microbiotic community which mirrors that of breastfed infants.
Data from the new multicentre PLATYPUS study have also highlighted the tolerance, safety, and growth effcacy of AAFs supplemented with HMOs in infants with moderate-to-severe CMPA.25 This multicentre, open-label, non-randomised, single-arm study evaluated AAF containing 2’FL and LNnT (Alfamino® HMO, Nestlé) in 32 infants aged 1–8 months over a 4-month period. Normal growth, as measured by for-age Z-scores for weight, length, and head circumference and compared with World Health Organization
(WHO) reference values, was achieved in infants fed the study formula with two HMOs. There was even some evidence of catch-up growth, noted Vandenplas. The study formula had an excellent safety profile and was tolerated well by infants with CMPA.25
Vandenplas concluded that, while breast milk remains the best option, the addition of two HMOs (2’FL and LNnT) to infant formula brings the second choice for infant feeding one step closer to this gold standard. Evidence supports the effcacy of HMOs added to w-EHF containing lactose with the additional clinical benefits of decreased respiratory tract infections and antibiotic use. Supplementation with these two HMOs also achieves normal
growth in infants with moderate to severe CMPA fed with an AAF formula.
Human Milk Oligosaccharides Shaping the Gut Microbiome in Infants with Cow’s Milk Protein Allergy
Ralf Heine
The early development of the gut microbiome in infancy can be divided into three stages that take place during the first 1,000 days of
life: developmental, transitional, and stable. Determinants of early microbiome development were evaluated in the TEDDY study, which analysed 12,500 sequential stool samples from 903 children from birth to 26 months.26 Several significant dietary and environmental factors were identified but breast milk had “the stand-out influence in the early years of life,” said Heine, explaining that the reason for this is the strong effect of HMOs on shaping the gut microbiome.26
2’FL in human milk promotes healthy bacteria such as bifidobacteria and lowers the growth of pathogenic strains like streptococci, as evidenced by a study of relative abundance in infants from secretor versus non-secretor mothers.27 This HMO is therefore a major modifier in bacterial composition in early infancy, stressed Heine. Overall, HMOs have a range of effects that help support the immune system of infants and young children with CMPA, including promoting the growth of beneficial bacteria. A study of cow’s milk-based infant formula in healthy infants supplemented with 2’FL and LNnT showed that the faecal microbial composition of infants fed the HMO supplemented formula was significantly different to control formula and shifted closer to that of breastfed infants, particularly regarding Bifidobacteriaceae abundance.24
Heine went on to review microbiome data from the multicentre CINNAMON study that evaluated the growth, tolerance, and safety of a w-EHF containing 2’FL and LNnT versus a virtually identical control formula with no HMOs in infants with CMPA.21-22,28 The per-protocol set for the microbiome analysis included 132 infants. Each group contained similar numbers of infants (68 control and 64 HMO), and the male:female ratio was similar. The HMO group was slightly younger than the control group (mean age at enrolment 98.6 days versus 107.9 days, respectively) but still in a comparable range.28 Stool collections were performed during the CINNAMON study at four timepoints: baseline (Visit [V] 0, with infants aged between 14 days and 6 months at enrolment), after 1 and 3 months follow-up (V1 and V3), and at 12 months of age (V6). Heine explained that, although there was a significant age gradient during early visits, at V6 all infants were synchronised at 12 months of age, allowing for a clearer assessment and comparison of microbiome outcomes at that timepoint. Gut microbiome composition analysis was carried out via metagenomic sequencing, analysing the microbial genetic information, and profiling the bacterial species accordingly. Microbial richness and diversity were compared between groups at each timepoint using the Shannon index. There was also an enrichment analysis at the genus, family, and phylum level, focusing mainly on bifidobacteria. The study tracked faecal community types (FCTs) over time in order to develop a transition model and analyse the temporal development of the microbiome.
Data from the CINNAMON study looking at the evolution of faecal microbial diversity over time from V0 to V6 showed that at 12 months of age (V6), the gut microbiome of infants receiving HMO-supplemented test formula had lower diversity, with significant differences in richness (p=0.003) and Shannon index (p=0.006).28 The effect of HMO supplementation on the abundance of certain bacterial species was also evaluated after 1 month at V1. When grouped by genus, there was a significant enrichment of Bifidobacterium species, with these key bacteria being more abundant in HMO-treated infants (p=0.0002).28
A post hoc subanalysis was also carried out in infants aged <3 months at V0 (i.e., before starting a complementary diet, as the introduction of fibre is itself is an important modifier of bacterial composition) to assess whether the change in Bifidobacterium abundance between V0 and V1 was affected by mode of delivery. The enrichment of Bifidobacterium in young infants fed HMO-supplemented formula was much more pronounced in those born via Caesarean section, although a non-significant trend for increased Bifidobacterium was also seen in vaginally delivered infants receiving HMO-supplemented formula. These results highlight that the beneficial effect of supplemental HMOs on Bifidobacterium was greatest in infants born via Caesarean section, suggested Heine, but is still found across the spectrum.28
Faecal communities within the gut microbiome were shown to evolve with age during the first year of life in both study groups in the CINNAMON study, reflecting changes in diet and environmental exposures. In infants fed either HMO-supplemented or control formula, transition models showed a temporal development from ‘early’ to ‘late’ FCT clusters, using all 481 study samples, reflecting a gradual change to an ‘older’ microbiome. Importantly, the positive effect of HMO supplementation on microbiome age was confirmed by comparing differences in FCT distribution between treatment groups, stratified by visit from enrolment to V6 (Figure 1). At 12 months, the microbiome of HMO-treated infants was shown to be significantly enriched in early-type FCT as compared with control (p=0.014).28 The progression to more advanced microbiome FCTs was more advanced in the control group, which, according to Heine, “represents an acceleration of the microbiome age towards an adult-type gut microbiome in non-breastfed infants.” Overall, the evidence demonstrates that HMOs play a central role in shaping the developing gut microbiome of infants with CMPA. Supplementation with 2’FL and LNnT in both standard and w-EHF formula is associated with an enrichment in bifidobacteria and lower microbial diversity, as well as a lower gut microbiota age at 12 months because of the effects of HMOs. HMO supplementation therefore appears to slow the premature shift towards an adult-type gut microbiome, which was previously described in infants receiving no/limited breast milk, Heine concluded. The long-term clinical benefits of HMOs on early immune development and the resolution of CMPA require further study.
Immunological and Microbial Effects of Human Milk Oligosaccharides in Experimental Models of Food Allergy
Sophie Nutten
Breastfeeding is currently recommended to prevent the development of allergic diseases in high-risk infants; however, the data are conflicting and underlying mechanisms of prevention remain unclear.29 Breast milk consists of essential nutrients plus numerous bioactive compounds that could theoretically impact immune development in the neonatal gut.29 Nutten explained that HMOs have been “heavily studied over the last decade” and are now recognised as a major bioactive component of breast milk that could modulate the development of allergic disease.


HMOs comprise the third largest solid component in breast milk after lipids and lactose. Every mother synthesises and secretes a distinct and individualised composition of HMOs influenced by various factors including maternal genetics, lactation stage, and environmental conditions.
HMOs are undigested so have no nutritive function but support the infant immune system in four main ways.30 They promote a bifidobacteria- dominated microbiome, deflect pathogens, and strengthen gut barrier function.31 Approximately 1% of HMOs are also absorbed into the systemic circulation and interact directly with immune cells to educate the developing immune system.30 For all these reasons, HMOs have been proposed to influence the development of allergic disease. This hypothesis has been supported by several observational studies in humans showing that HMO composition is associated with the development of food sensitisation and other types of allergies.6,11,31,32 No interventional study has yet been published, so the causality of these associations remains to be determined; however, pre-clinical data suggest a benefit of HMO supplementation in both allergy prevention and amelioration of allergic symptoms.
In the first pre-clinical study to demonstrate the protective effect of HMOs, addition of 2’FL or 6’-sialyllactose (6’SL) was shown to attenuate food allergy symptoms such as diarrhoea in a mouse model induced by oral ovalbumin challenge.33 The addition of HMOs was also associated with a reduction in mouse mast cell proteinase 1 in the serum, indicating less mast cell degranulation, and an increase in IL-10+CD4+CD25+ cell populations in Peyer’s patches and mesenteric lymph nodes.33 In a passive cutaneous anaphylaxis model, oral administration of 2’FL and 6’SL induced suppression of antigen- induced mast cell degranulation by CD4+CD25+ cells isolated from the mesenteric lymph nodes.33 Collectively, these data suggest that HMOs can reduce the symptoms of food allergy through induction of IL-10 regulatory cells and indirect stabilisation of mast cells, Nutten explained.
The second pre-clinical study looked at the oral administration of 3'-sialyllactose (3'-SL) in two different mouse models of atopic dermatitis (AD).34 3’SL reduced ear thickness (a proxy for AD symptoms) and produced dose-dependent decreases in mast cell number and IgE production in the serum, while simultaneously reducing levels of inflammatory and AD-related cytokines.34
Additionally, in vitro assessment revealed that 3’SL was able to prevent skin inflammation by directly inducing transforming TGF-β mediated regulatory T-cell (T-reg) differentiation. This study also looked at the effects of 3’SL on the mouse microbiome and showed Bifidobacterium levels, in particular B. bifidium, increased in a dose-dependent manner with oral administration. Rising levels of Bifidobacterium were associated with the ‘rescue’ of AD phenotypes, noted Nutten. Taken together, these findings indicate that 3’SL has therapeutic effects against AD by inducing T-reg differentiation, downregulating AD-related cytokines and increasing the Bifidobacterium population.
Nutten went on to describe unpublished pre-clinical results obtained at Nestlé Research looking at the immunological and microbiome effects of HMOs. In the first experimental model, mice were sensitised with a mix of ovalbumin and cholera toxin and dietary intervention was performed with a mixture of 2’FL and LNnT in a 2:1 ratio, administered at increasing doses (0.2%,1%, 5%, 10% of total diet). Results showed that oral administration of the blend of two HMOs decreased allergic sensitisation induced via the gut as shown by a dose-dependent decrease in anti-ovalbumin IgG. Consistent with other studies, a decrease in mast cell numbers in the gut and mouse mast cell proteinase 1 in the serum was also seen, together with an increase in T-reg count in the mesenteric lymph nodes. Analysis of the microbiome effect of the HMO mix showed modulation of the gut microbiota composition with increasing doses of HMO, together with a decrease in the richness of the microbiota. Looking at specific species, Nutten noted that there was an increase in Akkermansia, which is known for its role in strengthening the gut barrier, and Parabacteroides, which is one of the predominant species in the intestinal tract.
In a separate model of allergy, mice underwent epicutaneous sensitisation with Aspergillus fumigatus applied to the skin and then received the same nutritional supplementation of HMOs as previously described. This model confirmed that oral administration of the blend of 2’FL and LNnT decreased allergic sensitisation induced via the skin, as shown by decreases in specific IgG1 and cytokines involved in Type 2 immune responses, such as IL-4, in the serum. Modulation of the murine gut microbiota was also observed with oral administration of the 2’FL and LNnT mix. HMOs promoted the relative abundance of Bacteroides and led to decreasing levels of Bilophila, a potentially deleterious genus linked to various diseases. Finally, the HMO mix was also shown to induce an increase in propionate production in the caecum. This SCFA has been associated with a decreased risk of allergy in children.
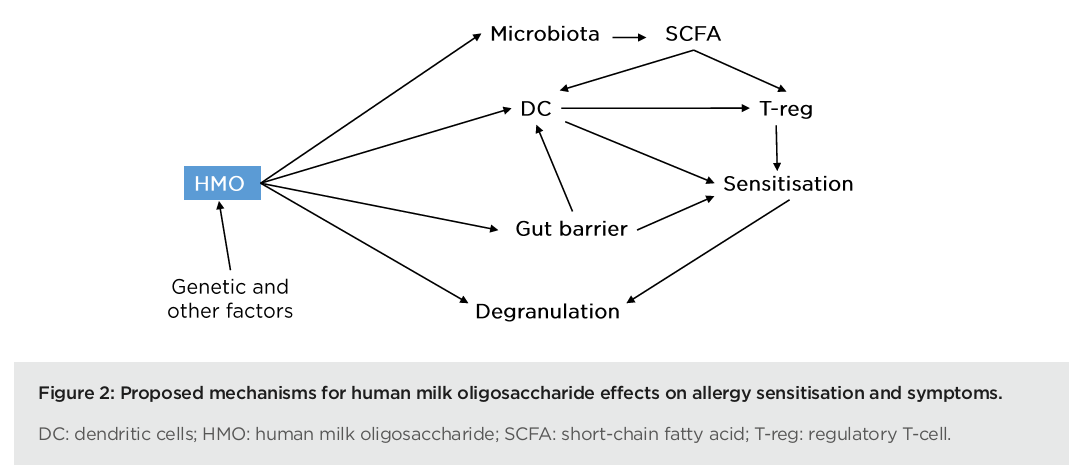
In summary, human observational studies suggest a key role of HMOs in allergy. Notably, HMO composition is associated with the development of food sensitisation in the first year(s) of life. However, human interventional studies are still needed to confirm the role of HMO supplementation in allergy management. Pre-clinical models have shown that supplementation of mice with HMOs has an impact on both allergy sensitisation and symptoms, and this research has also helped to decipher the underlying mechanisms of action (Figure 2). Data show that HMOs modulate the Type 2 immune response via interaction with immune cells such as basophils and dendritic cells. They also play a vital role in reinforcing the gut barrier and modulating the microbiota towards increased production of SCFAs, which promotes T-reg differentiation. These mechanisms could work together synergistically to explain the beneficial effects of HMOs in allergy, Nutten concluded.
Questions and Answers
As part of the interactive Question and Answer session that followed the main symposium, Vandenplas explained that HMOs protect against infection via several different mechanisms. HMOs stimulate the development of a healthy
microbiome by increasing bifidobacteria and lactobacilli and decreasing pathogenic strains; act as decoy receptors to block pathogen adherence to the gastrointestinal (GI) mucosa; and exert positive modulatory effects on the immune system. Asked what is known about risks in infants breastfed by non-secretor versus secretor mothers, Heine noted that the risk of infections (both GI and respiratory) and AD is reduced in secretors, suggesting that 2’-FL acts as an important immune modifier in this group to confer protective effects.
On the selection of particular HMOs for study in mouse models, Nutten explained that choice is driven by availability and the accumulated evidence from published association studies, which confirms an important role for 2’-FL. Nutten was also asked, from a mechanistic point of view, whether HMO effects in mice models are microbiome-dependent. She replied that the health benefits of HMOs are likely to arise from a combination of both microbiome-mediated effects and direct effects of HMOs on immune cells such as dendritic cells or basophils/mast cells.
In response to a question on the risks of microbiome deficiency in infants born by caesarean section, Heine noted that pure elective populations undergo inoculation with the ‘wrong’ bacteria (predominantly skin and environmental flora) and also tend to have lower rates of breastfeeding and increased antibiotic usage. All of these factors compound the risk of a bifidobacterial-deficient microbiome in the perinatal period, which could be corrected by using HMO-supplemented formulas if breastfeeding is not possible.
Looking at potential future areas of research, Vandenplas highlighted the important role of metabolites and the need for further study into specific HMOs of therapeutic relevance.
There is some literature to suggest differences in HMO profiles between allergic versus non-allergic children, he noted, but whether that is consequential or causal remains to be clarified.
A question from the audience queried the lack of dose-dependency in the final mouse study showcased by Nutten. She suggested this could indicate a plateau effect with 5% HMO, with the microbiota incapable of digesting higher concentrations. Another question from the audience raised the possibility of a role for HMOs in neural cross-talk in the developing neonatal gut nervous system. Vandenplas said this is currently unknown but it is clear that the microbiome is important in stimulating GI motility, with HMOs likely to play a role in this. There is also some data in healthy infants to show that probiotics result in more rapid gastric emptying.
The final question focused on the biological rationale for the natural diversity of HMOs seen in human breast milk. Heine explained that HMOs possess an incredibly complex array of functions and targeted effects, including a role in brain neurodevelopment. Moving forward, this space will continue to evolve, fuelled by improvements in the technical feasibility of making breast milk identical HMOs and increased understanding of their important immune and health benefits.
References
1. Stahl B et al. Oligosaccharides from human milk as revealed by matrix-
assisted laser desorption/ionization mass spectrometry. Anal Biochem.1994;223:218-26.
2. Field CJ et al. The immunological components of human milk and their
effect on immune development in infants. J Nutr. 2005;135:1-4.
3. Cederlund A et al. Lactose in human breast milk an inducer of innate
immunity with implications for a role in intestinal homeostasis. PLoS One.
2013;8:e53876.
4. Jalonen T. Identical intestinal permeability changes in children with different clinical manifestations of cow's milk allergy. JACI. 1991;88:737-42.
5. Tanaka M, Nakayama J. Development of the gut microbiota in infancy
and its impact on health in later life. Allergol Int. 2017;66(4):515-22.
6. Seppo AE et al. Human milk oligosaccharides and development of cow's milk allergy in infants. JACI.2017;139:708-11.
7. Van den Abbeele P et al. 2'-Fucosyllactose alters the composition and activity of gut microbiota from formula-fed infants receiving complementary feeding in a validated intestinal model. J Funct Foods. 2019;61:103484.
8. Austin S. Temporal change of the content of 10 oligosaccharides in
the milk of Chinese urban mothers. Nutrients. 2016;8(6):346.
9. Bode L. The functional biology of human milk oligosaccharides. Early Hum Dev. 2015;91(11):619-22.
10. Smilowitz J et al. Breast milk oligosaccharides: structure–function relationships in the neonate. Ann Rev Nutr. 2014;34(1):143-69.
11. Sprenger N et al. FUT2-dependent breast milk oligosaccharides and
allergy at 2 and 5 years of age in infants with high hereditary allergy risk. Eur J Nutr. 2017; 56(3):1293-301.
12. Björksten B et al. Allergy development and the intestinal microflora during the first year of life.J Allerg Clin Immunol. 2001;108:516-20.
13. Kalliomäki M et al. Distinct patterns of neonatal gut microflora in infants
in whom atopy was and was not developing. J Allerg Clin Immunol. 2001;107(1):129-34.
14. Woicka-Koljwa K et al. Food allergy is associated with recurrent respiratory tract infections during childhood. Postepy Dermatol Allergol. 2016;33(2):109 13.
15. Vandenplas Y. Prevention and management of cow’s milk allergy in non-exclusively breastfed infants. Nutrients. 2017;9(7):731.
16. Meyer R et al. When should infants with cow’s milk protein allergy use
an amino acid formula? A practical guide. J Allergy Clin Immunol Pract. 2018;6(2):383-99.
17. Nowak-Wegrzyn A et al. Confirmed hypoallergenicity of a novel whey-
based extensively hydrolyzed infant formula containing two human milk oligosaccharides. Nutrients. 2019;11(7):1447.
18. American Academy of Pediatrics. Committee on Nutrition. Hypoallergenic infant formulas. Pediatrics. 2000;106:346-9.
19. Urashima T et al. The predominance of type I oligosaccharides is a feature
specific to human breast milk. Adv Nutr. 2012;3:473S-82S.
20. Xu G et al. Absolute quantification of human milk oligosaccharides reveals phenotypic variations during lactation. J Nutr. 2017;147:117-24.
21. Vandenplas Y et al Growth tolerance and safety of an extensively hydrolyzed formula containing two human milk oligosaccharides in infants with cow’s milk protein allergy (CINNAMON Study). Abstract presented at EAACI-PAAM, Florence, Italy. October 17-19, 2019.
22. Vandenplas Y et al. Extensively hydrolysed formula with two human milk oligosaccharides reduces rate of upper respiratory tract infections in infants with cow’s milk allergy (CINNAMON study). Abstract. EAACI Digital, 6-8 June, 2020.
23. Puccio G et al. Effects of infant formula with human milk oligosaccharides on growth and morbidity: A randomized multicenter trial. J Pediatr Gastroenterol Nutr.2017;64:624-31.
24. Berger B et al. Linking human milk oligosaccharides, infant fecal community types, and later risk to require antibiotics. mBio. 2020;11:e03196-19.
25. Gold MS et al. Growth, tolerance and safety of an amino acid-based formula supplemented with two human milk oligosaccharides in infants with moderate-to-severe cow’s milk protein allergy. Abstract 628. EAACI Hybrid Congress, 10-12 July, 2021.
26. Stewart CJ et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature. 2018;562:583-8.
27. Lewis Z et al. Maternal fucosyltransferase 2 status affects the gut bifidobacterial communities of breastfed infants. Microbiome. 2015;
13):DOI: 10.1186/s40168-015-0071-z.
28. Pedersen H et al. An extensively hydrolysed formula supplemented
with two human milk oligosaccharides (HMO) shapes the gut microbiome in infants with cow’s milk protein allergy (CMPA). Abstract. FAAM-EUROBAT Digital 2020, 16-17 October, 2020.
29. Rajani PS et al. Immunologically active components in human milk and development of atopic disease, with emphasis on food allergy, in the pediatric population. Front Pediatr. 2018;6:218.
30. Rudloff S et al. Urinary excretion of in vivo ¹³C-labelled milk oligosaccharides in breastfed infants. Br J Nutr. 2012;107:957-63.
31. Miliku K et al. Human milk oligosaccharide profiles and food sensitization among infants in the CHILD Study. Allergy. 2018;73:2070-3.
32. Lodge CJ et al. Human milk oligosaccharide profiles and allergic disease up to 18 years. J Allergy Clin Immunol. 2021;147:1041-8.
33. Castillo-Courtade L et al. Attenuation of food allergy symptoms following
treatment with human milk oligosaccharides in a mouse model. Allergy. 2015;70:1091-102.
34. Li-Jung Kang et al. 3'-Sialyllactose prebiotics prevents skin inflammation
via regulatory T cell differentiation in atopic dermatitis mouse models. Scientific Reports. 2020;10:5603.
If you liked this post you may also like


