The protein intake of breastfed term infants has been used as the basis for estimating an infant’s protein requirements during the first year. Daily protein gain is highest in the very young infant and decreases rapidly in later infancy and in the second year of life. The protein content of breast milk evolves depending on the stage of lactation and time since delivery. Indeed, protein concentration in breast milk is high during the first few weeks of lactation and gradually subsides throughout the first year. The quantity and quality of breast milk is critical to support infant growth and long-term development.
Current knowledge
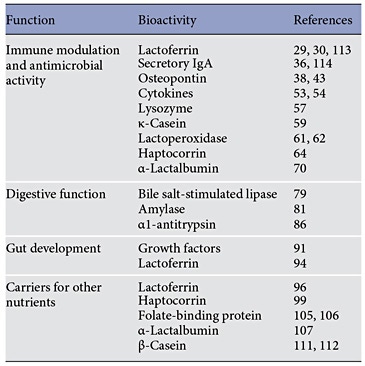
Proteins are the third most abundant solids found in breast milk. The variety of functions performed by the bioactive proteins and peptides in breast milk shed light on why breastfed infants have lower morbidity and fewer infections. Lactoferrin, secretory IgA, osteopontin, and various cytokines modulate the infant’s immune system alongside lysozyme, ĸ-casein, and lactoperoxidase, which have antibacterial functions. Other proteins regulate gut development and aid in the absorption of key nutrients.
Practical implications
Based on our better understanding of protein evolution in breastmilk across the stages of lactation, new infant formulas with lower protein concentration but better protein quality have been developed, tested, and made available in many countries. Low-birth-weight infants have higher protein requirements than term infants because of their higher daily protein gain per unit body weight. The concentrations of protein and amino acids in the breast milk of mothers who deliver preterm are higher during the first weeks of lactation compared to those of mothers who deliver at term. Supplementation of breast milk is needed to meet the high protein requirements of infants with very low and extremely low birth weight.
Recommended reading
Lönnerdal B, Erdmann P, Thakkar SK, Sauser J, Destaillats F: Longitudinal evolution of true protein, amino acids and bioactive proteins in breast milk: a developmental perspective. J Nutr Biochem 2016;41:1–11.
Protein intake of breastfed term infants has been used as a model to estimate protein requirements during the first year. They are higher during the first months when daily weight gain is fast and lower during later infancy when daily weight gain slows down.
Breast milk contains a multitude of bioactive proteins that are highly concentrated in early lactation and decrease with progressing lactation.
Quantity and quality of protein in breast milk are crucial for healthy growth and long-term development.
Protein ingested with breast milk provides indispensable amino acids which are necessary for new protein synthesis for growth and replacement of losses via urine, feces, and the skin. Protein gain in the body of an infant is highest during the first months when protein concentrations in breast milk are higher than during later stages of lactation. Low-birth-weight infants have higher protein needs than term infants and need protein supplements during feeding with breastmilk. Based on our better understanding of protein evolution in breastmilk during the stages of lactation, new infant formulas with lower protein concentration but better protein quality have been created, successfully tested, and are now available in many countries. Besides providing indispensable amino acids, bioactive protein in breast milk can be broadly classified into 4 major functions, that is, providing protection from microbial insults and immune protection, aiding in digestive functions, gut development, and being carriers for other nutrients. Individual proteins and their proposed bioactivities are summarized in this paper in brief. Indeed, some proteins like lactoferrin and sIgA have been extensively studied for their biological functions, whereas others may require more data in support to further validate their proposed functions.
Introduction
Breastfeeding is important for the healthy growth and development of infants and young children. The WHO recommends exclusive breastfeeding until 6 months and continuation of breastfeeding until 2 years as part of a mixed diet. However, recent DHS surveys indicate that even in developing countries only about 32% of mothers exclusively breastfeed their infants until 6 months [1], and the quality of complementary foods is very low. Therefore, in many developing countries, stunting is still prevalent in about 20% of children under 5 years of age [2]. In most developed countries, solids are introduced between 4 and 6 months, and breastfeeding is often stopped much earlier than recommended.
After carbohydrates and lipids, proteins are the third abundant solids in breast milk (BM), not only providing crucial amino acids indispensable for growth but also bioactive proteins and peptides essential for many functions (Table 1).
Table 1. Bioactive functions of breast milk proteins
Protein Needs for Growth
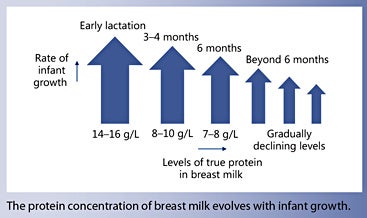
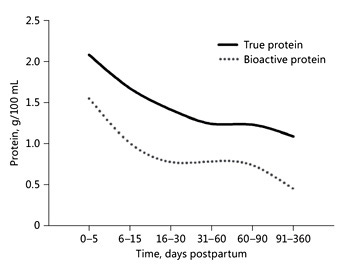
The protein intake of breastfed term infants has been used as a model to estimate protein requirements during the first year [3, 4]. The protein content in BM can be quantified by directly assessing the true protein content [5]. True protein concentrations of 14–16, 8–10, and 7–8 g/L have been reported during early lactation, at 3–4 months, and at 6 months, respectively. A recent metaanalysis of 43 studies [5] confirms that the protein concentration in BM depends on the stage of lactation and time since delivery. It also indicates a big variety in protein concentration, in particular during the first few months of lactation. However, the true protein intake does not accurately reflect the amount of utilizable amino acids to synthesize new body protein because some (bioactive) BM proteins can be found intact in infant stools [6].
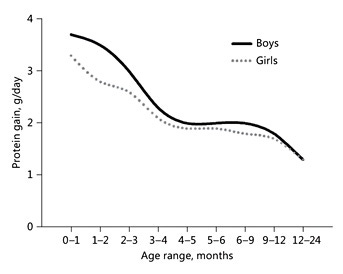
Protein requirements for growth and daily turnover strongly depend on rates of body protein gain. Daily protein retention in the body can be calculated by measuring absorption and excretion. Fomon et al. [7] published detailed estimates of protein gain in children during the first 2 years and beyond. Based on total body water (estimating the extra- and intracellular compartment), total body potassium (estimating the intracellular compartment where almost all of the protein is present), and total body calcium (osseous minerals), all components of fat-free mass were calculated. Because potassium is the main intracellular cation, gains at different age ranges allow to estimate gain of total body protein (Fig. 1). Daily protein gain is highest in the very young infant and is rapidly decreasing during later infancy and the second year of life: during the first months, protein gain is 3 times higher than between 12 and 24 months. Indeed, protein concentration in BM is high during the first few weeks of lactation and then continues to decrease throughout the first year, but at substantially lower rates than those observed in the first weeks (Fig. 2). Casein and most whey proteins in BM are utilized for growth. Their concentrations change profoundly over the course of lactation: during the first 2 weeks of lactation, concentrations of whey proteins are very high, while concentrations of caseins are low, which results in a whey:casein ratio as high as 80:20. The ratio drops to 65:35 by week 2 and stays constant at about 60:40 thereafter [5].
Fig. 1. Daily protein gains of boys and girls aged 0–24 months. Gains are based on total body potassium measurements [7].
Fig. 2. Evolution of true protein (solid line) and bioactive protein (dotted line; sum of lactoferrin, α-lactalbumin, serum albumin, sIgA, IgG, IgM, lysozyme, well-characterized bioactive proteins) of breast milk within the first year of postpartum life. Based on data from Lönnerdal et al. [5].
Proteins are polymer chains made of amino acids linked together by peptide bonds. During the digestion process, most proteins are decomposed to simple amino acids or small peptides which are absorbed. Amino acids which are absorbed and not oxidized are the building blocks of new protein which is synthesized in the body. Despite the reduction in protein over time, the nutritional value of protein in BM, as measured by the ratio of essential amino acids to total amino acids, appears to be consistent over time. These changes correlate well with the evolving needs of the growing infant [8].
Because of their higher daily protein gain per unit of body weight, low-birth-weight infants have higher protein requirements than term infants [9, 10]. Concentrations of protein and amino acids during the first weeks of lactation are higher in BM of mothers who deliver preterm than in BM of mothers who deliver at term [8]. However, feeding BM without supplements does not meet protein requirements, in particular those of verylow- and extremely-low-birth-weight infants. Supplements which are on the market are based on protein fractions of cow’s milk or human donor milk. Supplementation of BM or the use of preterm formulas improves growth rates of low-birth-weight infants, but it is questionable if the right mix of indispensable amino acids for synthesis of new body protein in low-birth-weight infants has already been found.
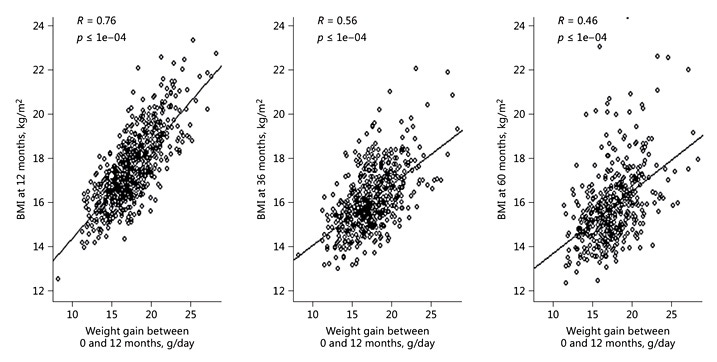
Growth differences between breastfed infants and infants fed high-protein formulas (i.e. >2.25 g/100 kcal) have been shown more than 2 decades ago [11]. Infants fed a formula with high protein content grow faster during the first 2 years and beyond [12], and they have higher insulin and insulin-like growth factor 1 levels in blood [13, 14]. Rapid growth during the first year is associated with obesity during childhood. Three longitudinal studies confirm a strong relationship between weight gain between 0 and 12 months and BMI at 12, 36, and 60 months (Fig. 3) [10]: 21% of the variance of BMI at 60 months was explained by weight gain between birth and 12 months. Narrowing the protein gap between BM and infant formula requires a deep understanding of how protein quality and quantity in BM changes over time. Composition of infant formulas has evolved with increasing knowledge of BM. Recently, a pooled analysis of individual growth data (11 clinical trials; n = 1,882 [15]) of infants who received a modified whey-based low-protein starter formula (1.8 g protein/100 kcal) [16] with an amino acid profile close to term BM [17] has become available. The weight and length of formula-fed infants at 4 months correspond to the 50th percentile of the WHO global growth standard [18] . The CHOP study in Europe followed the growth of children who had been fed low- or high-protein formulas during the first year of life. At 6 years, BMI and the percentage of children with obesity were significantly lower in the low-protein formula group [12]. Two randomized controlled trials [19, 20] tested 2 low-protein follow-up formulas (1.6 g protein/100 kcal) and followed children until 5 years of age. BMI at 5 years of the children who had been on the low-protein formula was similar to that of the children who had been exclusively breastfed until 4–6 months. [10].
Therefore, it seems that the quantity and quality of protein in BM are crucial for healthy growth and longterm development. Some nutritive proteins which are partly or well absorbed may also have biological functions. In addition, there are bioactive proteins in BM which are not absorbed. Specific functions of bioactive proteins and peptides which have been studied in detail can provide insights on why breastfed infants have lower morbidity and shorter infection periods [21] as well as different microbiota [22].
Fig. 3. Weight gain between birth and 12 months predicts BMI at 12, 36, and 60 months. Data from 3 longitudinal studies (Italy, USA, Chile) [10].
Bioactive (Functional) Proteins in BM
Immunomodulatory and Antimicrobial Activity
Lactoferrin Lactoferrin, also known as lactotransferrin, was reported to be present in bovine milk in the late 1930s and quantified in BM in the early 1960s [23]. Originally described as “red protein from (bovine) milk,” it turned out to be a multifunctional globular glycoprotein [24, 25]. Lactoferrin content of BM decreases with progressing stages of lactation, found highest in colostrum at 5.5 g/L and between 1.5 and 3.0 g/L in mature milk depending on the stage of lactation [26]. It is generally accepted that lactoferrin resists digestion to some degree and therefore can be found intact in infant feces. However, early in life, a fraction of this protein may be taken up by the intestinal mucosa and the rest is digested to yield bioactive peptides [27]. Due to its high affinity towards ferric iron, it not only acts as a carrier of iron in BM, but also deprives harmful microbes of iron that is key for their growth. Additionally, due to its basic N-terminal domain that can interact with various microbial and host targets, lactoferrin not only has antimicrobial properties but also modulates the innate and adaptive immune responses. This is orchestrated by cytokines interleukin (IL)-4, IL-2, IL-12, and interferon-γ and results in its ability to act as an antior a proinflammatory agent. It has been demonstrated that lactoferrin can compete with lipopolysaccharide for binding to CD14 and thereby preventing lipopolysaccharide- mediated proinflammatory series of events [28]. Although this molecule is resilient to digestion and can be found intact in fecal material, it is digested to some extent to form lactoferricin, a molecule that is able to inhibit Escherichia coli attachment to intestinal cells [29, 30]. Lactoferricin may not be the only lactoferrin related peptide to have antimicrobial activity, the role of Lf(1–11) and lactoferrampin has also emerged in the recent past [31]. In one study, Lf(1–11) was demonstrated to be active against gram-positive bacteria (Staphylococcus spp. and Streptococcus mitis) as well as gram-negative bacteria (Acinetobacter baumannii, Pseudomonas spp., Klebsiella spp., and E. coli) [32].
Secretory Immunoglobulin A Mothers’ milk is rich in secretory immunoglobulin A (sIgA) especially during early stages of lactation. Colostrum shows a wide variability from mother to mother and may contain on average 2.0 g/L of sIgA, which is reduced to approximately 0.5 g/L in mature milk [33]. The absorptive fate of this protein is similar to that of lactoferrin in that it is partly excreted intact and partly digested to bioactive peptides [27] . This class of antibody has been documented to be abundantly present in intestinal mucosa of humans and other mammals and to protect the epithelium from toxic assaults. As a first line of defense, in inhibiting infectious incident, sIgA would block toxin adhering to intestinal epithelium. In mouse models exposed to Vibrio cholerae toxin, sIgA demonstrated a protective effect [34]. Another mechanism by which sIgA could block pathogens is by direct recognition of receptor-binding domains like reovirus type 1 Lang. When IgA knockout mice were challenged with reovirus, the orally gavaged IgA group of knockout mice was as effective as wild-type mice in clearing the infection [35]. sIgA may also have a direct effect on the virulence of the bacteria. For example, murine monoclonal IgA-binding Shigella flexneri suppressed activity of the bacterial type 3 secretion (T3S), necessary for S. flexneri to gain entry into intestinal epithelium [36]. Immune exclusion is often referred to as sIgA’s ability to prevent pathogen access to intestinal epithelium through a series of processes involving agglutination, entrapment in mucus, and clearance via peristaltic movements [37].
Osteopontin Osteopontin is a multifunctional, heavily glycosylated and phosphorylated acidic protein with possible roles in immune activation, inhibition of ectopic calcification, cellular adhesion and migration, angiogenesis, and bone remodeling [38]. With high variability amongst populations and stages of lactation, the average concentration of osteopontin in BM is approximately 140 mg/L [39]. When compared to wild-type suckling mice, osteopontin knockout suckling mice suffered from prolonged periods and intense bouts of diarrhea upon rotavirus exposure [40]. A fine balance between T helper 1 (Th1) and T helper 2 (Th2) is required to alleviate an immune response. Osteopontin has been demonstrated to induce Th1 expression and inhibit Th2 along with IL-10 [38]. Interestingly, breastfed infants but not formula-fed infants showed induction of Th1 response when immunized with measles, mumps and rubella [41]. This observation can presumably be tied to presence of osteopontin in BM of the breast fed group but not the formula fed group. Furthermore, through electrostatic interactions, osteopontin can form complexes with lactoferrin and thereby acts as a carrier for other immunomodulatory proteins to further enhance the immune competency of its consumers [42]. Recently, a randomized controlled trial was carried out where 2 groups of formula-fed infants and a group of breast-fed infants were recruited. The 2 formula-fed groups were fed a standard formula with 65 mg/L bovinederived osteopontin or an experimental formula containing 130 mg/L bovine-derived osteopontin. Apart from comparable growth parameters, differences were observed in lowered levels of proinflammatory cytokine TNF-α and a significantly fewer number of days when the infants had fever [43].
Cytokines The effect of cytokines to regulate inflammatory processes often associated with infection is usually like an orchestra, operating in network and produces a cascading effect. Cytokines are postulated to enhance proliferation of thymocytes, inhibit IL-2 production from T-cells and suppress IgE production [44–46]. The presence of several cytokines in BM has been demonstrated over years. These molecules include, but may not be limited to, IL-1β, IL-6, IL-8, IL-10, TNF-α, interferon-γ, transforming growth factor-β, and colony-stimulating factor [47–52]. Usually they are present at very low concentrations (picograms) and potentially may originate from epithelial mammary gland cells, or activated macrophages and other cells in BM [53]. The biological function of these agents on infants is to complement infants’ own source of cytokines that are produced at lower quantities due to immaturity of the immune system. Although cytokines are not as well studied as other immunomodulatory agents described in this section, it is postulated that these molecules balance Th1 and Th2 to impart immunity-related benefits [54].
Lysozyme Lysozyme belongs to the whey class of protein fraction in BM and possesses bactericidal properties by affecting the cell wall of most of gram-positive and some gramnegative bacteria. Higher amounts of lysozyme have been observed in colostrum at approximately 0.36 g/L that is reduced slightly in mature milk to 0.30 g/L [55]. Attempts have also been made to produce recombinant human lysozyme and lactoferrin in dairy animals [56] . The mechanism is yet unclear, but lysozyme of BM origin also contains activity against HIV type 1 [57].
κ-Casein κ-Casein belongs to the casein family of phosphoproteins that is involved in a number of physiological processes. With an average concentration of approximately 1.25 g/L in colostrum and transition milk, it settles closer to 1 g/L in mature milk [58]. These glycosylated κ-casein of human origin as opposed to bovine origin inhibit the cell lineage-specific adhesion of Helicobacter pylori to human gastric surface mucous cells [59].
Lactoperoxidase A member of the heme peroxidase family, lactoperoxidase is secreted by mammary glands and is persistently present during lactation. In human BM, lactoperoxidase is found at 1–1.5 units/mL during the first 6 months of lactation [60]. It is well accepted that this peroxidase catalyzes oxidation of thiocyanate from the saliva of infants to hypothiocyanate in the presence of small amounts of hydrogen peroxides already in the mouth of the baby. The formed hypothiocyanate may be responsible to exterminate gram-positive and gram-negative bacteria [61, 62].
Haptocorrin Haptocorrin is a vitamin B 12 -binding protein found in many body fluids including BM with a concentration range of approximately 5 μg/ mL in colostrum to 3 μg/mL in mature milk [63]. Structurally, haptocorrin did not show much alternations after exposure to digestive enzymes and was able to inhibit the growth of E. coli in an in vitro system [64]. Further systematic study of exposure of haptocorrin to 34 commensal and pathogenic bacteria indicated suppression of only Bifidobacterium breve implicating its role that may be limited to certain strains and a blanket antimicrobial label might not be relevant for this protein warranting further studies [65].
α-Lactalbumin A well-characterized and primary protein in BM, α- lactalbumin is made up of 123 amino acids and 4 disulfide bridges and accounts for 20–25% of total BM proteins [66, 67]. Since it is also a rich source of many indispensable amino acids, a fraction of the protein is digested well and the rest yields polypeptides that exert antimicrobial activities mostly against gram-positive bacteria and not gram-negative bacteria [68, 69]. Additionally, a folding variant of α-lactalbumin was also found to be bactericidal against an antibiotic resistant strain of Streptococcus pneumonia [70]. Not only for its antimicrobial benefits but also to mimic the BM closer for additional benefits, all efforts have been made to enrich BM substitute with α-lactalbumin [71].
Digestive Functions
Bile Salt-Stimulated Lipase The major source of energy for breast-fed infants is the predominant form of lipid in BM, the triacylglycerols. Milk triacylglycerols are efficiently digested by complementary actions of gastric lipase, colipase-dependent pancreatic lipase, and bile salt-stimulated lipase (BSSL). While there are 2 sources of these enzymes, infants’ exocrine pancreas, the major source is maternal milk BSSL. In the early 1950s, it was first demonstrated by Freudenberg that mothers’ milk contains an inactive lipase that is stimulated when the chyme reaches duodenum and comes in contact with bile salts [72, 73]. BSSL was purified and characterized in the early 1980s [74], is demonstrated to have a broad substrate specificity [75–77] and to inactivate by pasteurization of BM [78]. Therefore, digestion and absorption of lipids is significantly lower when pasteurized donor milk is fed to preterm infants [79]. Recently in a randomized phase 3 study, recombinant human BSSL was added to infant formula to assess if it had any impact on growth velocity, presumably due to better lipid digestion and absorption. Interestingly, the benefits on growth were not observed in appropriate-forgestational- age preterm infants but were present in smallfor- gestational-age preterm infants [80].
Amylase In absence of pancreatic amylase, BM amylase may catalyze hydrolysis of starch, glycogen and other related saccharides by cleavage of α-1,4 linkages to produce maltose, dextrins, and glucose. Activity of amylase varies from 1,000 to 5,000 units per liter of BM [81]. Colostrum is known to contain higher activities compared to transition or mature milk [82]. There is a further decrease of approximately 35% of the activity beyond the first trimester of breastfeeding [83]. Additionally, higher parity may also reduce amylase activity by half [83]. Preterm milk contains equal amounts of amylase activity as term milk [84]. Apart from its obvious digestion-aiding role, amylase may also act as antibacterial by attacking the polysaccharides of the bacterial cell wall [82].
α1-antitrypsin Thy physiological role of protease inhibitors like α1- antitrypsin (A1AT) in BM is not completely understood. However, as generally accepted for other mammals, protease inhibitors may play a role in digestion and/or absorption of bioactive proteins present at relatively higher concentrations in colostrum. Indeed, McGilligan et al. [85] showed the highest presence of A1AT in colostrum (1.4–5.2 g/L) compared to the first 26 weeks (0.07 g/L) or 26–52 weeks of lactation (0.05 g/L). A1AT resists digestion in the enteral tract and can be found intact in feces of infants in significant quantities [86]. Efforts have been made to express human A1AT in transgenic sheep for potential human applications [87, 88].
Gut Development
Growth Factors Growth factors, their concentrations in BM, and biological sources of growth factors have been described in the literature [89]. Potentially originating from epithelial and stromal cells as well as from macrophages of mammary glands, the growth factors are present at microgram-per-liter quantities in BM. Growth factors that are present in the intestinal lumen, such as epidermal growth factor and insulin-like growth factors 1 and 2, originate either from salivary glands of the infants or from mothers’ milk [90]. It still remains unclear how they are able to exert effects upon their receptors that are located on the basolateral side of the absorptive intestinal epithelial cells. Indeed, one suggestion is that the immature gut of the infant provides access of the ligands to the basolateral compartment. Preterm infants, whose gut is relatively underdeveloped compared to that of term infants, may have significantly higher concentrations of epidermal growth factor in milk secreted by their mothers [91].
Lactoferrin Lactoferrin exposure to intestinal cell culture models shows increased proliferation and differentiation in a dose-dependent manner [92] . Additionally, it also has greater proliferation of intestinal crypt cells in a piglet model [93] . Indeed, it is plausible that rapid maturation of absorptive intestinal epithelia in presence of lactoferrin may contribute to the higher weight gain in infants fed BM substitute with lactoferrin compared to a control group without lactoferrin [94].
Carriers for Other Nutrients
Lactoferrin Iron absorption in breastfed infant was reported to be more efficient than cow’s milk-based infant formula [95]. Certainly, this was later attributed to thepresence of relatively high levels of lactoferrin in BM compared to bovine milk (approx. 1 mg/mL and 10 μg/mL, respectively) and the majority of the iron in milk is bound to lactoferrin [25, 26]. Furthermore, a receptor of lactoferrin was later identified that had greater affinity for human lactoferrin in contrast to bovine lactoferrin for iron absorption [96]. Efforts are underway to express recombinant human lactoferrin in rice, and a comparison of the stability and bioactivity has shown promising results for its potential use in BM substitutes [97].
Haptocorrin Haptocorrin, also known as transcobalamin 1, perhaps a name derived from “transporter of cobalamin,” an alternative name for vitamin B 12, is a binding protein found in BM [98]. In adults, vitamin B 12 absorption is dependent on digestive juices, enzymes, binding protein secreted in stomach, intrinsic factors, and their receptors in the small intestine [99]. However, in infants, very low amounts of intrinsic factors have been found in fecal material, perhaps indicating that haptocorrin may have a larger role to play in the transport of vitamin B 12 [100].
Folate-Binding Protein Identified in the late 1960s, folate-binding protein (FBP) almost entirely binds all naturally occurring folate in BM as well as bovine milk [101, 102]. Since its discovery, it has been thought that FBP sequesters various forms of folate and ensures adequate supply to the neonate by also preventing the oxidation in the digestive tract [103, 104]. Solids of pooled human and bovine milk contained approximately 2,000 nmol/kg, whereas goat milk contains twice as much and freeze-drying or spray-drying of milk to powder retains practically all FBP [105, 106]. Since FBP is able to withstand digestion, it is plausible that permeable intestinal lining of the infant gut is able to take up the folate-FBP complex at least for weeks or even months postpartum until the tight junctions are formed [54].
α-Lactalbumin Originally proposed to carry divalent cations like calcium and zinc [54], it did show higher absorption in infant rhesus monkeys [107]. However, research work in human infants did not show any signs of altered absorption of iron α-lactalbumin-enriched infant formula [108], warranting further studies for this biological benefit.
β-Casein The total casein concentration increases during lactation and represents approximately 10–20% during earlier stages and 40–50% when lactation matures [109, 110]. In mature milk, β-casein may represent up to 25% or approximately 2.7 g/L in BM [58]. This protein is highly phosphorylated, which at least in preclinical model has shown to solubilize calcium and uptake by intestinal cells at least in part by forming casein phosphopeptides that may act as calcium ionophores or calcium carriers across the membrane [111, 112]. More research remains to be done to elucidate the role of casein phosphopeptides in enhanced uptake of other divalent cations like zinc and even iron.
Conclusion
Recent findings on nutritive and bioactive proteins in breastmilk support the WHO recommendations that breastfeeding should be continued during the first year and beyond. Infant formula manufacturers should eliminate all high-protein formulas from the market. New formulas for infants should be low in protein, in particular follow-up formulas and growing-up milks. Protein quality in formulas (amino acid profiles) should be closer to that in BM. Before bioactive proteins are added to infant formulas, safety and efficacy tests must be provided by formula manufacturers.
Disclosure Statement
F.H. is a board member of the Nestlé Nutrition Institute, a nonprofit- making Swiss association which receives educational grants from Nestec S.A., Switzerland, and other sources. S.K.T. is an employee of Nestlé Research Center, Lausanne, Switzerland.
References
Haschke F, et al: Feeding patterns during the first 2 years and health outcome. Ann Nutr Metab 2013; 62(suppl 3):16–25.
Global Burden of Disease Study 2013 Collaborators: Global, regional, and national incidence, prevalence, and years lived with disability for 301 acute and chronic diseases and injuries in 188 countries, 1990–2013: a systematic analysis for the Global Burden of Disease Study 2013. Lancet 2015; 386: 743– 800.
Fomon SJ: Requirements and recommended dietary intakes of protein during infancy. Pediatr Res 1991; 30: 391–395.
Heinig MJ, et al: Energy and protein intakes of breast-fed and formula-fed infants during the first year of life and their association with growth velocity: the DARLING Study. Am J Clin Nutr 1993; 58: 152–161.
Lönnerdal B, Erdmann P, Thakkar SK, Sauser J, Destaillats F: Longitudinal evolution of true protein, amino acids and bioactive proteins in breast milk: a developmental perspective. J Nutr Biochem 2016; 41: 1–11.
Donovan SM, et al: Partition of nitrogen intake and excretion in low-birth-weight infants. Am J Dis Child 1989; 143: 1485–1491.
Fomon SJ, et al: Body composition of reference children from birth to age 10 years. Am J Clin Nutr 1982; 35(suppl):1169–1175.
Zhang Z, et al: Amino acid profiles in term and preterm human milk through lactation: a systematic review. Nutrients 2013; 5: 4800– 4821.
Ziegler EE, et al: Body composition of the reference fetus. Growth 1976; 40: 329–341.
Haschke F, Grathwohl D, Haiden N: Metabolic programming: effects of early nutrition on growth, metabolism and body composition. Nestle Nutr Inst Workshop Ser 2016; 86: 87–95.
Dewey KG, et al: Breast-fed infants are leaner than formula-fed infants at 1 y of age: the DARLING study. Am J Clin Nutr 1993; 57: 140–145.
Weber M, et al: Lower protein content in infant formula reduces BMI and obesity risk at school age: follow-up of a randomized trial. Am J Clin Nutr 2014; 99: 1041–1051.
Axelsson IE, Ivarsson SA, Raiha NC: Protein intake in early infancy: effects on plasma amino acid concentrations, insulin metabolism, and growth. Pediatr Res 1989; 26: 614– 617.
Socha P, et al: Milk protein intake, the metabolic- endocrine response, and growth in infancy: data from a randomized clinical trial. Am J Clin Nutr 2011; 94(suppl):1776S–1784S.
Alexander DD, et al: Growth of infants consuming whey-predominant term infant formulas with a protein content of 1.8 g/100 kcal: a multicenter pooled analysis of individual participant data. Am J Clin Nutr 2016; 104: 1083–1092.
Raiha NC, et al: Whey predominant, whey modified infant formula with protein/energy ratio of 1.8 g/100 kcal: adequate and safe for term infants from birth to four months. J Pediatr Gastroenterol Nutr 2002; 35: 275– 281.
Haschke F, et al: Postnatal high protein intake can contribute to accelerated weight gain of infants and increased obesity risk. Nestle Nutr Inst Workshop Ser 2016; 85: 101– 109.
WHO: WHO Child Growth Standards: Length/Height-for-Age, Weight-for-Age, Weight-for-Length, Weight-for-Height and Body Mass Index-for-Age, Methods and Development. Geneva, WHO, 2016.
Inostroza J, et al: Low-protein formula slows weight gain in infants of overweight mothers. J Pediatr Gastroenterol Nutr 2014; 59: 70– 77.
Ziegler EE, et al: Adequacy of infant formula with protein content of 1.6 g/100 kcal for infants between 3 and 12 months. J Pediatr Gastroenterol Nutr 2015; 61: 596–603.
Dewey KG, Heinig MJ, Nommsen-Rivers LA: Differences in morbidity between breast-fed and formula-fed infants. J Pediatr 1995; 126: 696–702.
Isolauri E: Development of healthy gut microbiota early in life. J Paediatr Child Health 2012; 48(suppl 3):1–6.
Blanc B, Bujard E, Mauron J: The amino acid composition of human and bovine lactotransferrins. Experientia 1963; 19: 299–301.
Siqueiros-Cendon T, et al: Immunomodulatory effects of lactoferrin. Acta Pharmacol Sin 2014; 35: 557–566.
Lonnerdal B, Iyer S: Lactoferrin: molecular structure and biological function. Annu Rev Nutr 1995; 15: 93–110.
Rai D, et al: Longitudinal changes in lactoferrin concentrations in human milk: a global systematic review. Crit Rev Food Sci Nutr 2014; 54: 1539–1547.
Lonnerdal B: Bioactive proteins in breast milk. J Paediatr Child Health 2013; 49(suppl 1):1–7.
Actor JK, Hwang SA, Kruzel ML: Lactoferrin as a natural immune modulator. Curr Pharm Des 2009; 15: 1956–1973.
Tomita M, et al: Potent antibacterial peptides generated by pepsin digestion of bovine lactoferrin. J Dairy Sci 1991; 74: 4137–4142.
Edde L, et al: Lactoferrin protects neonatal rats from gut-related systemic infection. Am J Physiol Gastrointest Liver Physiol 2001; 281:G1140–G1150.
Bruni N, et al: Antimicrobial activity of lactoferrin- related peptides and applications in human and veterinary medicine. Molecules 2016; 21:E752.
Brouwer CP, Rahman M, Welling MM: Discovery and development of a synthetic peptide derived from lactoferrin for clinical use. Peptides 2011; 32: 1953–1963.
Goldman AS, et al: Immunologic factors in human milk during the first year of lactation. J Pediatr 1982; 100: 563–567.
Lycke N, et al: Lack of J chain inhibits the transport of gut IgA and abrogates the development of intestinal antitoxic protection. J Immunol 1999; 163: 913–919.
Silvey KJ, et al: Role of immunoglobulin A in protection against reovirus entry into Murine Peyer’s patches. J Virol 2001; 75: 10870– 10879.
Forbes SJ, et al: Transient suppression of Shigella flexneri type 3 secretion by a protective O-antigen-specific monoclonal IgA. MBio 2011; 2:e00042-11.
Stokes CR, Soothill JF, Turner MW: Immune exclusion is a function of IgA. Nature 1975; 255: 745–746.
Ashkar S, et al: Eta-1 (osteopontin): an early component of type-1 (cell-mediated) immunity. Science 2000; 287: 860–864.
Schack L, et al: Considerable variation in the concentration of osteopontin in human milk, bovine milk, and infant formulas. J Dairy Sci 2009; 92: 5378–5385.
Maeno Y, et al: Effect of osteopontin on diarrhea duration and innate immunity in suckling mice infected with a murine rotavirus. Viral Immunol 2009; 22: 139–144.
Pabst HF, et al: Differential modulation of the immune response by breast- or formulafeeding of infants. Acta Paediatr 1997; 86: 1291–1297.
Azuma N, Maeta A, Fukuchi K, Kanno C: A rapid method for purifying osteopontin from bovine milk and interaction between osteopontin and other milk proteins. Int Dairy J 2006; 16: 370–378.
Lonnerdal B, et al: Growth, nutrition, and cytokine response of breast-fed infants and infants fed formula with added bovine osteopontin. J Pediatr Gastroenterol Nutr 2016; 62: 650–657.
Soder O: Isolation of interleukin-1 from human milk. Int Arch Allergy Appl Immunol 1987; 83: 19–23.
Hooton JW, et al: Human colostrum contains an activity that inhibits the production of IL-2. Clin Exp Immunol 1991; 86: 520–524.
Sarfati M, et al: Presence of IgE suppressor factors in human colostrum. Eur J Immunol 1986; 16: 1005–1008.
Munoz C, et al: Interleukin-1 beta in human colostrum. Res Immunol 1990; 141: 505–513.
Rudloff HE, et al: Tumor necrosis factor-alpha in human milk. Pediatr Res 1992; 31: 29– 33.
Saito S, et al: Detection of IL-6 in human milk and its involvement in IgA production. J Reprod Immunol 1991; 20: 267–276.
Garofalo R, et al: Interleukin-10 in human milk. Pediatr Res 1995; 37: 444–449.
Eglinton BA, Roberton DM, Cummins AG: Phenotype of T cells, their soluble receptor levels, and cytokine profile of human breast milk. Immunol Cell Biol 1994; 72: 306–313.
Grosvenor CE, Picciano MF, Baumrucker CR: Hormones and growth factors in milk. Endocr Rev 1993; 14: 710–728.
Lonnerdal B: Human milk proteins: key components for the biological activity of human milk. Adv Exp Med Biol 2004; 554: 11– 25.
Lonnerdal B: Nutritional and physiologic significance of human milk proteins. Am J Clin Nutr 2003; 77: 1537S–1543S.
Golinelli LP, Del Aguila EM, Flosi Paschoalin VM, Silva JT, Conte-Junior CA: Functional aspect of colostrum and whey proteins in human milk. J Hum Nutr Food Sci 2014; 2: 1035.
Cooper CA, Maga EA, Murray JD: Production of human lactoferrin and lysozyme in the milk of transgenic dairy animals: past, present, and future. Transgenic Res 2015; 24: 605–614.
Lee-Huang S, et al: Lysozyme and RNases as anti-HIV components in beta-core preparations of human chorionic gonadotropin. Proc Natl Acad Sci USA 1999; 96: 2678–2681.
Cuilliere ML, et al: Changes in the kappacasein and beta-casein concentrations in human milk during lactation. J Clin Lab Anal 1999; 13: 213–218.
Stromqvist M, et al: Human milk kappa-casein and inhibition of Helicobacter pylori adhesion to human gastric mucosa. J Pediatr Gastroenterol Nutr 1995; 21: 288–296.
Shin K, Tomita M, Lonnerdal B: Identification of lactoperoxidase in mature human milk. J Nutr Biochem 2000; 11: 94–102.
Bjorck L, et al: Antibacterial activity of the lactoperoxidase system in milk against pseudomonads and other gram-negative bacteria. Appl Microbiol 1975; 30: 199–204.
Aoki,Y, Kobayashi K, Kajii T: Enzyme-linked immunoassay of haptocorrin: analysis of milk and granulocytes. Biochem Med Metab Biol 1992; 47: 189–194.
Adkins Y, Lonnerdal B: Potential host-defense role of a human milk vitamin B- 12-binding protein, haptocorrin, in the gastrointestinal tract of breastfed infants, as assessed with porcine haptocorrin in vitro. Am J Clin Nutr 2003; 77: 1234–1240.
Jensen HR, et al: Effect of the vitamin B12- binding protein haptocorrin present in human milk on a panel of commensal and pathogenic bacteria. BMC Res Notes 2011; 4: 208.
Lonnerdal B, Lien EL: Nutritional and physiologic significance of alpha-lactalbumin in infants. Nutr Rev 2003; 61: 295–305.
Permyakov EA, Berliner LJ: Alpha-lactalbumin: structure and function. FEBS Lett 2000; 473: 269–274.
Pellegrini A, et al: Isolation and identification of three bactericidal domains in the bovine alpha-lactalbumin molecule. Biochim Biophys Acta 1999; 1426: 439–448.
Wada Y, Lonnerdal B: Bioactive peptides derived from human milk proteins – mechanisms of action. J Nutr Biochem 2014; 25: 503–514.
Hakansson A, et al: A folding variant of alpha- lactalbumin with bactericidal activity against Streptococcus pneumoniae . Mol Microbiol 2000; 35: 589–600.
Lien EL: Infant formulas with increased concentrations of alpha-lactalbumin. Am J Clin Nutr 2003; 77: 1555S–1558S.
Blackberg L, et al: The bile salt-stimulated lipase in human milk is an evolutionary newcomer derived from a non-milk protein. FEBS Lett 1980; 112: 51–54.
Freudenberg E: Lipase of human milk; studies on its enzymological and nutritional significance. Bibl Paediatr 1953; 9: 1–68.
Blackberg L, Hernell O: The bile-salt-stimulated lipase in human milk. Purification and characterization. Eur J Biochem 1981; 116: 221–225.
Blackberg L, et al: Bile salt-stimulated lipase in human milk and carboxyl ester hydrolase in pancreatic juice: are they identical enzymes? FEBS Lett 1981; 136: 284–288.
Hernell O, Blackberg L: Digestion of human milk lipids: physiologic significance of sn-2 monoacylglycerol hydrolysis by bile saltstimulated lipase. Pediatr Res 1982; 16: 882– 885.
Lindquist S, Hernell O: Lipid digestion and absorption in early life: an update. Curr Opin Clin Nutr Metab Care 2010; 13: 314– 320.
Fredrikzon B, et al: Bile salt-stimulated lipase in human milk: evidence of activity in vivo and of a role in the digestion of milk retinol esters. Pediatr Res 1978; 12: 1048– 1052.
Andersson Y, et al: Pasteurization of mother’s own milk reduces fat absorption and growth in preterm infants. Acta Paediatr 2007; 96: 1445–1449.
Casper C, et al: Recombinant bile salt-stimulated lipase in preterm infant feeding: a randomized phase 3 study. PLoS One 2016; 11:e0156071.
Heitlinger LA, et al: Mammary amylase: a possible alternate pathway of carbohydrate digestion in infancy. Pediatr Res 1983; 17: 15– 18.
Lindberg T, Skude G: Amylase in human milk. Pediatrics 1982; 70: 235–238.
Dewit O, Dibba B, Prentice A: Breast-milk amylase activity in English and Gambian mothers: effects of prolonged lactation, maternal parity, and individual variations. Pediatr Res 1990; 28: 502–506.
Hegardt P, et al: Amylase in human milk from mothers of preterm and term infants. J Pediatr Gastroenterol Nutr 1984; 3: 563–566.
McGilligan KM, Thomas DW, Eckhert CD: Alpha-1-antitrypsin concentration in human milk. Pediatr Res 1987; 22: 268–270.
Davidson LA, Lonnerdal B: Fecal alpha 1-antitrypsin in breast-fed infants is derived from human milk and is not indicative of enteric protein loss. Acta Paediatr Scand 1990; 79: 137–141.
Wright G, et al: High level expression of active human alpha-1-antitrypsin in the milk of transgenic sheep. Biotechnology (NY) 1991; 9: 830–834.
Carver A, et al: Expression of human alpha 1 antitrypsin in transgenic sheep. Cytotechnology 1992; 9: 77–84.
Donovan SM, Odle J: Growth factors in milk as mediators of infant development. Annu Rev Nutr 1994; 14: 147–167.
Playford RJ, Macdonald CE, Johnson WS: Colostrum and milk-derived peptide growth factors for the treatment of gastrointestinal disorders. Am J Clin Nutr 2000; 72: 5–14.
Dvorak B, et al: Increased epidermal growth factor levels in human milk of mothers with extremely premature infants. Pediatr Res 2003; 54: 15–19.
Buccigrossi V, et al: Lactoferrin induces concentration-dependent functional modulation of intestinal proliferation and differentiation. Pediatr Res 2007; 61: 410–414.
Reznikov EA, et al: Dietary bovine lactoferrin increases intestinal cell proliferation in neonatal piglets. J Nutr 2014; 144: 1401– 1408.
Hernell O, Lonnerdal B: Iron status of infants fed low-iron formula: no effect of added bovine lactoferrin or nucleotides. Am J Clin Nutr 2002; 76: 858–864.
Saarinen UM, Siimes MA, Dallman PR: Iron absorption in infants: high bioavailability of breast milk iron as indicated by the extrinsic tag method of iron absorption and by the concentration of serum ferritin. J Pediatr 1977; 91: 36–39.
Kawakami H, Lonnerdal B: Isolation and function of a receptor for human lactoferrin in human fetal intestinal brush-border membranes. Am J Physiol 1991; 261:G841– G846.
Suzuki YA, et al: Expression, characterization, and biologic activity of recombinant human lactoferrin in rice. J Pediatr Gastroenterol Nutr 2003; 36: 190–199.
Burger RL, Allen RH: Characterization of vitamin B12-binding proteins isolated from human milk and saliva by affinity chromatography. J Biol Chem 1974; 249: 7220–7227.
Seetharam B, Alpers DH: Absorption and transport of cobalamin (vitamin B12). Annu Rev Nutr 1982; 2: 343–369.
Adkins Y, Lonnerdal B: Mechanisms of vitamin B(12) absorption in breast-fed infants. J Pediatr Gastroenterol Nutr 2002; 35: 192–198.
Nygren-Babol L, Jagerstad M: Folate-binding protein in milk: a review of biochemistry, physiology, and analytical methods. Crit Rev Food Sci Nutr 2012; 52: 410–425.
Ford JE, Salter DN, Scott KJ: A folate-protein complex in cow’s milk. Proc Nutr Soc 1969; 28: 39A–40A.
Ford JE: Observations on the possible nutritional significance of vitamin-binding proteins in milk. Proc Nutr Soc 1974; 33: 15A.
Ford JE: Some observations on the possible nutritional significance of vitamin B12-and folate-binding proteins in milk. Br J Nutr 1974; 31: 243–257.
Wigertz K, Svensson UK, Jagerstad M: Folate and folate-binding protein content in dairy products. J Dairy Res 1997; 64: 239– 252.
Indyk HE, Filonzi EL, Gapper LW: Determination of minor proteins of bovine milk and colostrum by optical biosensor analysis. J AOAC Int 2006; 89: 898–902.
Kelleher SL, et al: Glycomacropeptide and alpha-lactalbumin supplementation of infant formula affects growth and nutritional status in infant rhesus monkeys. Am J Clin Nutr 2003; 77: 1261–1268.
Szymlek-Gay EA, et al: α-Lactalbumin and casein-glycomacropeptide do not affect iron absorption from formula in healthy term infants. J Nutr 2012; 142: 1226–1231.
Kunz C, Lonnerdal B: Re-evaluation of the whey protein/casein ratio of human milk. Acta Paediatr 1992; 81: 107–112.
Kunz C, Lonnerdal B: Human-milk proteins: analysis of casein and casein subunits by anion-exchange chromatography, gel electrophoresis, and specific staining methods. Am J Clin Nutr 1990; 51: 37–46.
Sato R, Noguchi T, Naito H: Casein phosphopeptide (CPP) enhances calcium absorption from the ligated segment of rat small intestine. J Nutr Sci Vitaminol (Tokyo) 1986; 32: 67–76.
Ferraretto A, et al: Casein phosphopeptides influence calcium uptake by cultured human intestinal HT-29 tumor cells. J Nutr 2001; 131: 1655–1661.
Liu KY, et al: Natural killer cell populations and cytotoxic activity in pigs fed mother’s milk, formula, or formula supplemented with bovine lactoferrin. Pediatr Res 2013; 74: 402–407.
Hurley WL, Theil PK: Perspectives on immunoglobulins in colostrum and milk. Nutrients 2011; 3: 442–474.