The lipid components of human milk provide the infant with energy and essential micronutrients, and they also serve specific roles to support gastrointestinal function, lipid and lipoprotein metabolism, neurodevelopment, and immunity. There have been significant advances both in food technology, which enables the supply of new lipid preparations, and in lipidomic analyses, which offer insight into the biological effects of complex lipids in infancy. These will pave the way for improvements in the feeding of infants who cannot be breastfed.
Current knowledge
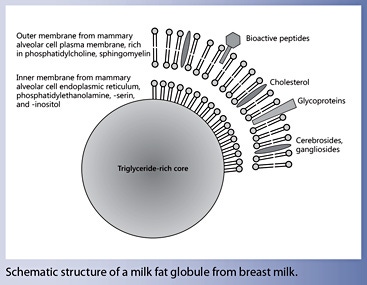
Human milk lipids provide a major portion of the total energy intake in infants (approximately half of the energy supply). The concentration of milk lipids varies greatly between individuals, during the day, and throughout the course of breastfeeding. The hindmilk contains a higher fat composition and a correspondingly larger mean size of the milk fat globule. The outer layer of the milk fat globule membrane (MFGM) consists of a bilayer of amphipathic lipids, primarily phosphatidylcholine, sphingomyelin, and cholesterol, as well as cerebrosides, gangliosides, and others. These components are highly bioactive.
Practical implications
The biological importance of MFGM is gaining increased attention after several clinical trials reported benefits of adding components of MFGM to infant formula. Current evidence supports the provision of omega-3 docosahexaenoic acid along with omega-6 arachidonic acid with infant formula. The recent revision of the European legislation that was implemented in 2016 stipulates that all infant and follow-on formula must contain between 20 and 50 mg omega-3 docosahexaenoic acid per 100 kcal without a minimum requirement of arachidonic acid. This is a novel concept never clinically tested for suitability and safety of healthy infants from birth, and indications of possible adverse effects exist. Therefore, we recommend not to use such formula until conclusive data on their safety might become available in the future.
Recommended reading
Grote V, Verduci E, Scaglioni S, Vecchi F, Contarini G, Giovannini M, et al: Breast milk composition and infant nutrient intakes during the first 12 months of life. Eur J Clin Nutr 2016;70:250– 256.
Human milk lipids provide a major portion of the energy supply to breastfed infants as well as essential vitamins, polyunsaturated fatty acids, complex lipids, and bioactive components.
Recent data evaluating the addition of preparations of complex lipids with or without milk fat globule membranes to vegetable oil-based infant formula show promising indications for potential improvements of infant development and reduction of infection risk.
Analyses of gene-diet interaction following the concept of Mendelian randomization add to the evidence that the supply of long-chain polyunsaturated fatty acids in infancy is causally related to improving cognitive development and to reducing asthma risk at school age. Current evidence supports the provision of omega-3 docosahexaenoic acid along with omega-6 arachidonic acid with infant formula.
Significant methodological progress both in food technology enabling the provision of new lipid preparations and in lipidomic analyses offers major opportunities to explore the biological effects of the supply of complex human milk lipids.
Human milk lipids provide the infant with energy and essential vitamins, polyunsaturated fatty acids, and bioactive components. Adding complex lipids and milk fat globule membranes to vegetable oil-based infant formula has the potential to enhance infant development and reduce infections. Cholesterol provision with breastfeeding modulates infant sterol metabolism and may induce long-term benefits. Some 98–99% of milk lipids are comprised by triacylglycerols, whose properties depend on incorporated fatty acids. Attention has been devoted to the roles of the longchain polyunsaturated fatty acids docosahexaenoic (DHA) and arachidonic (ARA) acids. Recent studies on gene-diet interaction (Mendelian randomization) show that breastfeeding providing DHA and ARA improves cognitive development and reduces asthma risk at school age particularly in those children with a genetically determined lower activity of DHA and ARA synthesis. It appears prudent to follow the biological model of human milk in the design of infant formula as far as feasible, unless conclusive evidence for the suitability and safety of other choices is available. The recent European Union legislative stipulation of a high formula DHA content without required ARA deviates from this concept, and such a novel formula composition has not been adequately evaluated. Great future opportunities arise with significant methodological progress for example in lipidomic analyses and their bioinformatic evaluation, which should enhance understanding of the biology of human milk lipids. Such knowledge might lead to improved dietary advice to lactating mothers as well as to further opportunities to enhance infant formula composition.
Introduction
Lipids are a major source of energy provided with human milk to the infant [1, 2], but they also provide essential nutrients such as polyunsaturated fatty acids (PUFA) and lipid soluble vitamins. Many studies have demonstrated important biological effects of the milk lipids provided to the recipient infant, for example on gastrointestinal function, lipid and lipoprotein metabolism, membrane composition and function, infant growth, neurodevelopment, and immune function [3].
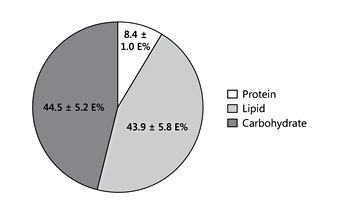
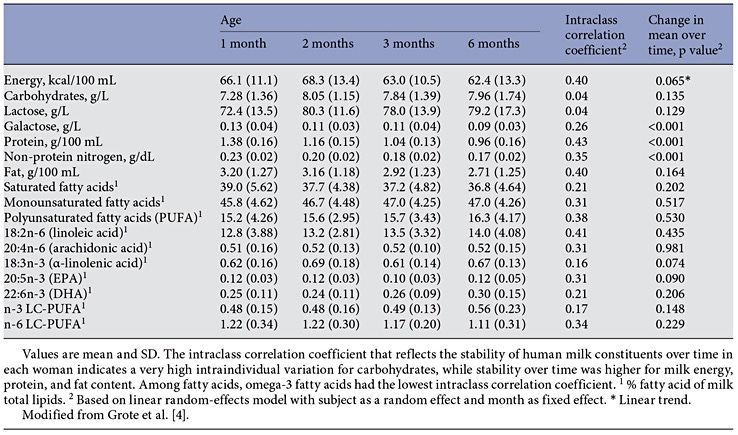
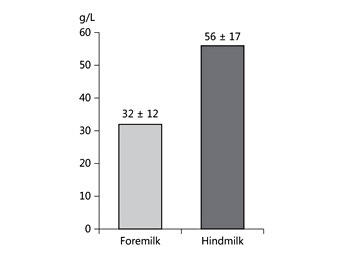
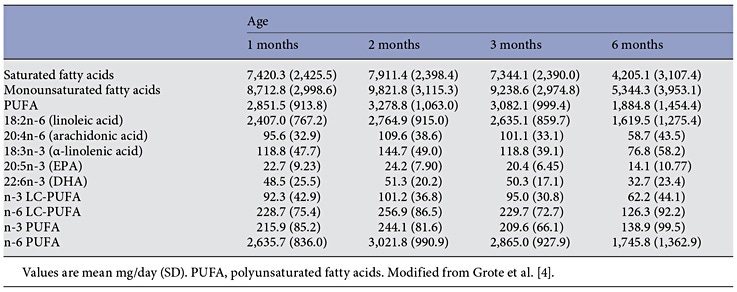
Human milk lipids provide a major portion of the total energy intake in young infants, with a mean 44% of the energy supply [4] (Fig. 1). The average intake of human milk lipids in fully breastfed infants amounts to 21.42 g/ day between birth and 6 months of age [4]. This results in an impressive 3.9 kg of human lipid supplied during the first half year of life to fully breastfed infants, equivalent to some 35,000 kcal provided by human milk lipids alone during the first 6 months of life. While the mean lipid content in human milk is relatively stable during the course of the first months of lactation, there is very wide interindividual and intraindividual variation of milk fat concentrations (Table 1) [4–6]. In fact, among the macronutrients in milk, fat shows the most variable concentration. For example, in mature milk samples collected at the infant age of 2 months, we find a coefficient of variation of 37.3% for milk fat but only of 14.4% for lactose and 12.9% for protein [4]. Milk fat content tends to increase with longer duration of breastfeeding and varies during the course of a day [1, 6]. Milk fat concentration increases with an increasing time interval to the preceding milk expression from the same breast, and it increases with maternal fat deposition in pregnancy indicated by the degree of gestational weight gain [7]. Milk fat increases during the course of each breastfeeding meal, with markedly higher milk fat contents in hindmilk (at the end of feeding) than in foremilk (at the beginning of the feed) (Fig. 2) [8]. This may be of biological benefit in that infants will initially get milk rich in the essential water-soluble substrates, whereas those who are hungrier and drink more milk obtain milk with an increasing fat and energy content to satisfy their caloric needs. Of interest, the increase of milk fat content during the meal is accompanied with a marked increase in the mean size of milk fat globule. Thereby, hindmilk has a higher ratio of triglycerides in the core of the milk fat globule (providing energy) to the surface membranes (rich in phospholipids, complex lipids, and essential long-chain polyunsaturated fatty acids, LC-PUFA).
Fig. 1. Contribution of macronutrients to total energy intake in breastfed infants aged 1 month. Drawn from data of Grote et al. [4] . E%, percetage of energy supply.
Table 1. Longitudinal evolution of human milk constituents in 30 prospectively followed lactating women
Fig. 2. Milk fat concentration in fore- and hindmilk collected before and after breastfeeding of 15 term infants. Drawn from data of Khan et al. [79] .
Milk Fat Globules and Complex Lipids
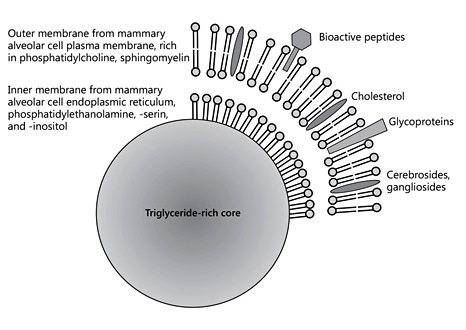
Milk can be characterized as an emulsion of milk fat globules in an aqueous liquid. Milk fat globules with markedly variable sizes are formed in the mammary alveolar cells and contain a core of nonpolar lipids comprised primarily of triacylglycerols, with added small amounts of monoglycerides, diglycerides, and nonesterified fatty acids. These nonpolar lipids are formed in the endoplasmic reticulum from fatty acids obtained from the maternal circulation as well as primarily intermediate- chain fatty acids with 12 and 14 carbon atoms synthesized from acetyl-CoA. Upon the secretion from the endoplasmic reticulum of mammary epithelial cells into the cytosol, this triglyceride-rich core is covered by an inner membrane derived from the endoplasmic reticulum consisting of a monolayer primarily of phosphatidylethanolamine, phosphatidylserine, phosphatidylinositol, and cholesterol. When these lipid droplets are further excreted from mammary epithelial cells into the alveolar space, they are covered by a piece of the apical plasma membrane, which results in the addition of another phospholipid bilayer and hence a phospholipid trilayer, and the other components of the mammary epithelial cell membrane such as membrane proteins and glycoproteins (Fig. 3). This outer layer of the milk fat globule membrane (MFGM) consists of a bilayer of amphipathic lipids, primarily phosphatidylcholine, sphingomyelin, and cholesterol, as well cerebrosides, gangliosides, glycosylated proteins and polypeptides, filaments, mucins, lactadherin, butyrophilin, and others; hence, MFGM contain a high density of bioactive components [9].
Fig. 3. Schematic depiction of human milk fat globules.
Phospholipids, plasmalogens, and sphingolipids including ceramides and gangliosides provide about 0.2– 1% of total milk lipids or about 100–400 mg/L [2]. The concentration of different phospholipids per 100 g milk were reported as 8.5 mg sphingomyelin, 6.8 mg phosphatidylethanolamine, 6.0 mg phosphatidylcholine, 1.4 mg phosphatidylserine, and 1.1 mg/100 g for phosphatidylinositol [10]. Phospholipids serve structural roles as indispensable components of all plasma membranes of body cells and organelles, and they have an impact on membrane functions and metabolism. Complex lipids also have roles in signal transmission and cell recognition [2, 3]. Gangliosides contribute some 10% of brain lipids, with high concentrations in the cerebral cortex.
The biological importance of MFGM is getting increased attention after several controlled trials reported benefits of adding bovine MFGM of complex lipid fractions to infant formula with fat derived predominantly from vegetable oil. A trial on formula enriched with sphingomyelin in preterm infants reported neurobehavioral benefits [11]. In a small trial in Indonesia, the addition of a ganglioside-rich bovine milk lipid fraction was reported to improve the hand and eye coordination IQ, performance IQ, and total IQ assessed with the Griffiths Mental Developmental Scale at the age of 24 weeks [12].
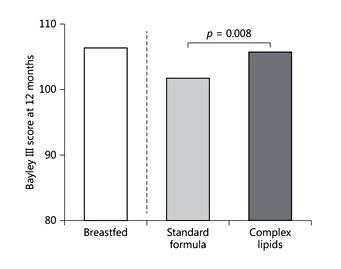
Another trial providing a milk formula with addition of a similar preparation for 12 weeks enrolled 450 infants aged 8–24 months in India and reported no difference for rotavirus or for all-cause diarrhea. In a large study that enrolled more than 500 Peruvian infants, MFGM-supplemented formula did not affect diarrhea incidence but reduced longitudinal diarrhea prevalence [13]. A larger trial that enrolled more than 250 toddlers aged 2.5–6 years in Belgium reported that a milk preparation enriched with a phospholipid-rich lipid fraction resulted in less days with fewer and lower parental scoring of internal, external, and total behavioral problems [14]. A further trial enrolled 160 formula-fed infants in Sweden as well as a breastfed reference group and evaluated effects of added bovine MFGM, along with reduced formula contents of energy and protein. The MFGM group achieved higher cognition scores in the Bayley test at the age of 1 year (Fig. 4) and showed a much lower incidence of acute otitis media as well as less use of antipyretic drugs [15, 16]. These observations lead to the conclusion that MFGM and/or the complex lipids provided with the MFGM fraction may have important biological roles for the development of nervous and immune functions.
Table 1. Infants fed a vegetable oil formula with an added bovine milk fat globule preparation with complex lipids and bioactive proteins showed an improved cognitive outcome at the age of 1 year compared to those fed standard formula, and were more similar to the test results in a breastfed reference group. Drawn from data of Timby et al. [16].
Cholesterol
Milk fat globule lipids also provide considerable amounts of free and esterified cholesterol, resulting in a total cholesterol content of 90–150 mg/L in human milk in contrast to typically only 0–4 mg/L in infant formula. Cholesterol is an indispensable building block for all cell membranes and is incorporated in considerable amounts into myelin in the nervous system during the period of rapid brain growth, and it serves as the substrate for the synthesis of bile acids, lipoproteins, vitamin D, hormones, and oxysterols that modulate cholesterol, lipid, and glucose homeostasis [3, 9, 17–19]. The provision of cholesterol with breastfeeding is associated with higher plasma concentrations of total and low-density lipoprotein cholesterol in breastfed than in formula-fed infants [20]. The provision of preformed cholesterol is most likely the cause for the about 3-fold lower endogenous cholesterol synthesis rate in breastfed than formula-fed infants, since the synthesis rate is inversely correlated to the daily cholesterol supply in mg/kg bodyweight [21]. In formula-fed piglets, dietary cholesterol supply downregulated hepatic hydroxymethylglutaryl coenzyme A reductase, the rate regulating enzyme for endogenous cholesterol synthesis [22]. In human infants aged 4 months, the rate of endogenous cholesterol synthesis also appeared to be regulated by dietary cholesterol supply. Breastfed infants with a high cholesterol and low phytoestrogen intake had the lowest fractional synthesis rate, while infants receiving cows’ milk-based formula with low cholesterol and low phytoestrogen had an intermediate rate, and infants fed soy-based formula with no cholesterol and high phytoestrogen had the highest rate of synthesis [23]. When cholesterol was added to the soy-based infant formula, the rate of synthesis was changed to similar results as in infants fed cows’ milk-based formula, which leads to the conclusion that the amount of dietary cholesterol supply regulates cholesterol synthesis in infants. Lasting effects of early feeding on later cholesterol levels were reported in several studies and reviewed in meta-analyses. A rather modest lowering of total and low-density lipoprotein cholesterol was found in adults who had been breastfed in infancy, compared to previously formula-fed people, with a greater effect size of exclusive than of partial breastfeeding [24, 25]. It was proposed that if 30% of infants are exclusively breastfed, resulting in a blood cholesterol reduction in adulthood by 0.15 mmol/L, the population prevalence of cardiovascular disease could be reduced by as much as 5% [25]. However, Ip et al. [26] noted that the analysis reporting reduced serum lipid levels in previously breastfed adults did not segregate the data according to gender and did not explicitly analyze potential confounders; they concluded that in view of the limited methodological quality of the meta-analysis the relationship between breastfeeding and adult cholesterol levels cannot be correctly characterized. Meta-analyses of available data do not allow definitive conclusions regarding the relationship between breastfeeding and all-cause mortality from cardiovascular diseases in adult life, although the confidence limits around the point estimates and the observed between-study heterogeneity do not exclude potential beneficial or adverse cardiovascular effects of breastfeeding [26, 27] . Therefore, it appears particularly promising to evaluate the short- and long-term effects of addition of well bioavailable preparations of cholesterol to infant formula in randomized controlled trials, which may shed further light on the potential biological importance of a dietary cholesterol supply in infancy.
Fatty Acids Provided with Milk Lipids
Triacylglycerols contribute some 98–99% of human milk fat. The properties of milk triglycerides are very much influenced by their fatty acid composition. Milk lipids of European women today typically contain some 35–40% saturated fatty acids, 45–50% monounsaturated fatty acids, and approximately 15% PUFA (Table 2). The saturated palmitic acid (C16: 0) provides approximately 25% of all milk fatty acids and hence the major part of the total saturated fatty acid content. About 70% of human milk palmitic acid is esterified in the middle position (sn- 2 position) of triacylglycerols which facilitates absorption. During intestinal digestion, fatty acids in the sn-1 and sn-2 positions are liberated as nonesterified fatty acids by pancreatic lipases. These nonesterified fatty acids are quite well absorbed if they are unsaturated and hence better water soluble. In contrast, liberated long-chain saturated fatty acids, such as palmitic acid, are poorly water soluble and poorly absorbed, but rather bind to calcium and form calcium soaps that are excreted with stools, thereby reducing both fat and calcium absorption. However, if palmitic acid is esterified in the sn-2 position, as it is predominantly the case in human milk lipids, pancreatic lipolysis yields a palmitoyl-monogylcerol which is well water soluble and well absorbed, thereby reducing fat and calcium malabsorption [28].
Table 2. Absolute fatty acid supply with human milk in prospectively followed lactating women
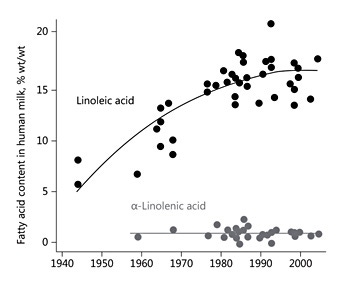
The human milk contents of the mono-unsaturated fatty acid oleic acid (C18: 1n-9) and of the essential PUFA linoleic acid (C18: 2n-6) and α-linolenic acid (C18: 3n-3) vary with the maternal dietary intake of these fatty acids. This is illustrated by the approximately 3-fold increase of linoleic acid content in mature human milk in the USA since the mid 1940s, along with the increase of dietary vegetable oil and linoleic acid consumption in the population, whereas α-linolenic acid contents have remained rather constant (Fig. 5) [29]. Thereby the average ratio of the omega-6 linoleic acid to the omega-3 α-linolenic acid in human milk has also increased approximately 3-fold. We studied the transfer of linoleic acid provided to lactating women into their milk using stable isotope-labelled fatty acids. An oral dose of 1 mg/kg bodyweight of linoleic acid uniformly labelled with the stable carbon isotope 13 C was provided repeatedly during the 2nd, 6th, and 12th week of lactation [30]. Before and at several times during a 5-day period after tracer intake, samples of breath and milk were collected, the volume of daily milk production was assessed, and dietary nutrient intakes were calculated from prospective dietary protocols. Some 3.5–4.5% of the ingested linoleic acid was oxidized to CO 2 and exhaled with breath, with no significant differences between the studied time points. Dietary linoleic acid was rapidly transferred into milk, with a peak enrichment reached about 12 h after intake (Fig. 6). Linoleic transfer into milk in unchanged form or as its metabolites did not change during the course of lactation. The data indicate that about 30% of milk linoleic acid is derived directly from dietary intake, whereas about 70% originates from maternal body fat stores. It is tempting to speculate that this largely indirect transfer of dietary linoleic via intermediate body stores may represent a biological benefit to the breastfed infant, since this mechanism buffers short-term variation of maternal dietary supply of the parent essential fatty acid and provides the infant with a relatively stable parent essential fatty acid supply. However, long-term changes in dietary supply will also modify maternal body fat stores and thereby explain the observed marked changes over time (Fig. 5). Only about 11% of the milk content of the linoleic acid metabolite dihomo-γ-linolenic acid (C20: 3n-6) in milk originates from direct endogenous conversion of maternal dietary linoleic acid, while only 1.2% of the milk arachidonic acid (ARA, C20: 4n-6) is directly derived from maternal linoleic acid intake [30].
Fig. 5. Evolution of the linoleic and α-linolenic acid contents in mature human milk in the USA over time. Drawn from data of Ailhaud et al. [29].
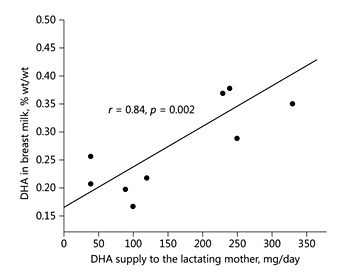
Fig. 6. The DHA supply to lactating women determines the DHA content in their breastmilk. Drawn from data of Fidler et al. [33].
Long-Chain Polyunsaturated Fatty Acids
The provision of LC-PUFA with milk, in particular of omega-6 ARA and omega-3 docosahexaenoic acid (DHA), has received considerable attention, because many of the biological effects of the essential fatty acids in early life appear to be mediated by LC-PUFA rather than the precursor essential fatty acids. Brenna et al. [31] performed a systematic review on 106 studies of human breast milk worldwide and culled to include only studies that used modern analysis methods capable of making accurate estimates of fatty acid contents as well as criteria related to the completeness of reporting. The final analysis included 65 studies with milk of 2,474 women. The authors found a milk ARA content of 0.47 ± 0.13% (mean ± SD, % wt/ wt), whereas milk DHA content was lower at 0.32 ± 0.22% [31]. Higher milk DHA contents were found in coastal populations and those with regular marine food consumption. The greater stability of milk ARA levels with a coefficient of variation of only 29%, as compared to DHA with a coefficient of variation of 69%, appears to reflect a greater degree of metabolic regulation of milk ARA content. Stable isotope studies have led us to the conclusion that 90% of human milk ARA are not derived directly from absorbed dietary lipids but rather from maternal ARA body stores [32]. In contrast, dietary DHA supply is a key determinant of milk DHA content. We showed that the dietary DHA intake is linearly correlated to milk DHA [33] (Fig. 6). Breastfeeding women need to achieve a daily DHA intake of at least 200 mg to provide milk with a DHA content of at least 0.3%, which is required for a fully breastfed infant to obtain the daily supply of about 100 mg DHA/day considered desirable to meet metabolic needs [34]. Given that the regulation of human milk ARA and DHA content differs, milk DHA and ARA are not closely correlated (r = 0.25, p = 0.02) [31], and the ARA/DHA ratio is not constant. It remains controversial whether the ratio of ARA to DHA in milk – or rather the amounts of DHA and of ARA supplied – are of greater relevance for biological effects in the infant. A balanced supply of both ARA and DHA appears to be relevant for the adequate incorporation of ARA and DHA into the growing brain [35].
In view of the marked accretion of ARA and DHA in the growing brain and the ample experimental evidence of the impact of LC-PUFA on membrane function, eicosanoid and docosanoid formation and the resulting regulation of physiological processes as well as the development and function of neural and immune tissues, the impact of LC-PUFA provision with human milk and also with infant formula has received considerable interest.
The provision of DHA was shown to enhance the early development of visual acuity. The European Food Safety Authority (EFSA) concluded that a cause and effect relationship has been established between the intake of infant and follow-on formula supplemented with DHA at levels around 0.3% of total fatty acids and visual function at 12 months in formula-fed infants born at term from birth up to 12 months and in breastfed infants after weaning up to 12 months [36]. However, some controversy remains with regards to the effects of the supply of preformed LC-PUFA on neurodevelopment of healthy term infants. For example, the authors of a meta-analysis on randomized trials evaluating infant formula with LCPUFA compared to formula without LC-PUFA concluded that while some studies showed a significant benefit, overall no significant effect was detectable [37, 38]. The authors noted the limitation of their conclusions by a large degree of heterogeneity of the included studies, which provided markedly different interventions and also used a variety of very different outcomes and approaches to outcome assessment. Of importance, the included studies did not adjust for the major impact of genetic variation modulating the rate of endogenous synthesis of LC-PUFA and related clinical endpoints, in particular variation in the Fatty Acid Desaturase (FADS) gene cluster [39, 40]. The lack of adjusting for this major modulating confounding factor in the included studies may considerably reduce the sensitivity to detect effects of dietary LC-PUFA effects. The comparison of breastfed infants provided with preformed LC-PUFA with infants fed formula without LC-PUFA in observational studies is also difficult to interpret, because human milk LC-PUFA and particularly DHA supply are closely associated with different dietary and lifestyle choices, including maternal smoking and parental socioeconomic status, which may also influence neurodevelopmental outcomes.
Further insight into PUFA effects are offered by considering the interaction of breastfeeding, which always supplies preformed LC-PUFA, and the genetic variation in the FADS gene cluster that predicts the enzyme activities of fatty acid desaturases 1 and 2. Gene variants of the FADS gene cluster have a major impact on the fatty acid composition of blood, tissues, and human milk [39–41] . We assessed the single-nucleotide polymorphisms in the FADS genes along with human milk fatty acid composition in 772 breastfeeding mothers who participated in the prospective Ulm Birth Cohort study both at 1.5 months after infant birth and at 6 months postpartum in a subset of 463 mothers who were still breastfeeding at this time [42]. At both time points, we found significant associations of FADS genotype with milk ARA contents and the ratio of ARA to dihomo-γ-linolenic acid, indicating that maternal FADS genotypes have an impact on the formation of LC-PUFA provided with breastmilk [42]. The variation of FADS genotypes was shown to also modulate the interaction of breastfeeding and cognitive development. Genotyping for the rs174575 variant in the FADS2 gene was performed in 5,934 children participating in the ALSPAC study in whom IQ tests had been performed at the age of about 8 years [43]. In line with other observational studies, previously breastfed children had higher IQ scores than previously formula fed children, but the relative impact of human milk nutrient supply and of confounding factors associated with breastfeeding cannot be easily deciphered from these observational data alone. Causal inferences on the role of human milk LCPUFA supply can be drawn from the fact that the beneficial effect of breastfeeding was much more pronounced, with an added advantage of about 4.5 IQ points, in the group of children with a genotype predicting a low ability of LC-PUFA synthesis [43]. Replication of these findings was published with the analysis of data from 2 Spanish birth cohort studies [44]. Since the genotype is considered to be distributed in the population at random (“Mendelian randomization”) and unrelated to the parental decision to breastfeed and to other related lifestyle predictors of IQ at school age, these data provide powerful evidence for the causality between early LC-PUFA supply and status during the breastfeeding period and later IQ achievements.
Galectins are important for cell turnover and immune regulation. The CRD of galectins is specific for β-gala ctosides. When cells are desialylated, the density of exposed galactose moieties on the cell surface increases. For example, naïve T cells express CD45 with an α-2,6-linked sialic acid. The amount of α-2,6-linked sialic acid is reduced following T-cell activation. The decrease in α-2,6- linked sialic acid renders the activated T cells susceptible to galectin-1-mediated apoptosis [48]. Thus, binding of sialyated HMO to cells may prevent apoptosis.
The relevance of LC-PUFA supply for child neurodevelopment was also demonstrated in a randomized clinical trial that enrolled 119 breastfeeding women in Texas, USA [45]. The women were assigned to receive identical capsules containing either a high-DHA algal oil providing approximately 200 mg DHA daily or a vegetable oil without DHA from delivery until 4 months after birth. Provision of DHA to the mother increased DHA in milk by about 70%, and in infant plasma phospholipids by about 20% [45]. At the age of 30 months, child psychomotor development was significantly better if mothers had received added DHA during the first 4 months of breastfeeding. At the age of 5 years, there were no differences in visual function, but children whose mothers had received added DHA performed significantly better on the Sustained Attention Subscale of the Leiter International Performance Scale (46.5 ± 8.9 vs. 41.9 ± 9.3, p < 0.008). These results support the conclusion that the DHA supply during early infancy is of importance for specific aspects of neurodevelopment.
Mendelian randomization also provided strong support for the conclusion that the LC-PUFA supply with breastfeeding is causally linked to protection against a later manifestation of bronchial asthma. Many studies have reported a protective effect of breastfeeding on asthma development, even though results are not consistent [26]. We evaluated the influence of the FADS1 FADS2 gene cluster polymorphisms on the association between breastfeeding and asthma in 2,245 children participating in 2 prospective German birth cohort studies, the GINI and LISA studies [46]. Logistic regression modelling was used to analyze the association between exclusive breastfeeding and doctor-diagnosed asthma occurring up to the age of 10 years, stratified by genotype. In the stratified analyses, heterozygous and homozygous carriers of the minor allele that show a low activity of LC-PUFA synthesis had a much reduced risk for later asthma if they were breastfed for 3 or 4 months and hence were provided with preformed LC-PUFA that can compensate for low endogenous synthesis (adjusted odds ratio between 0.37 [95% CI: 0.18–0.80] and 0.42 [95% CI: 0.20–0.88]). Interaction terms of breastfeeding with genotype were significant and ranged from –1.17 (p = 0.015) to –1.33 (p = 0.0066). Similarly, heterozygous and homozygous carriers of the minor allele who were exclusively breastfed for 5 or 6 months after birth had a reduced risk of asthma (0.32 [0.18–0.57] to 0.47 [0.27–0.81]) in the stratified analyses. In contrast, in individuals carrying the homozygous major allele predicting a greater degree of endogenous LC-PUFA formation, breastfeeding with provision of LC-PUFA showed no significant effect on asthma development. These results of a Mendelian randomization study demonstrate a lasting causal protection of breastfeeding for at least 3 months against doctor-diagnosed asthma until school age in children with a low rate of LC-PUFA synthesis and a modulating effect of postnatal PUFA status.
A systematic review on human studies on roles of LCPUFA and an expert workshop that reviewed the information and developed recommendations was recently performed with support from the Early Nutrition Academy [34]. It was concluded that breastfeeding women should get ≥ 200 mg DHA/day to achieve a human milk DHA content of at least ≈ 0.3% of fatty acids. Infant formula for term infants should contain DHA and ARA to provide 100 mg DHA/day and 140 mg ARA/day, and a supply of 100 mg DHA/day should continue during the second half of infancy. No quantitative advice on ARA levels in follow-on formula fed after the introduction of complimentary feeding was provided due to lack of sufficient data and considerable variation in ARA amounts provided with complimentary foods.
Should Infant Formula LC-PUFA Composition Be Guided by Human Milk Composition?
With regards to infant and follow-on formula, the recent revision of the European legislation that came into force in 2016 stipulates that all infant and follow-on formula must contain between 20 and 50 mg DHA/100 kcal (approx. 0.5–1% of fatty acids), whereas formula without DHA content will not be allowed any more to be placed on the European Union market once this legislation is implemented [47]. To the great surprise of many pediatricians and of experts in the field, no requirement for a minimum content of ARA in infant formula has been defined. This legal regulation is based on advice provided by the European Food Safety Authority that reviewed a variety of aspects and nutrients, including LC-PUFA DHA and ARA. In a first report on nutrient requirements and dietary intakes of infants and young children published in 2013, adequate LC-PUFA intakes were defined as 100 mg DHA/day and 140 mg ARA/day from birth to the age of 6 months, while 100 mg DHA/day were considered adequate from 6 to 24 months [48]. These conclusions are in line with many other scientific reports, including the recent recommendations of the Early Nutrition Academy- supported global expert group that are based on a systematic review of the available scientific evidence [34]. In contrast, the subsequently published EFSA report on the compositional requirements of infant and follow-on formula advised that all infant and follow-on formula should contain relatively high amounts of DHA at 20–50 mg/100 kcal, but without the need to provide any preformed ARA [49]. This DHA level stipulated by EFSA and the new European legislation is much higher than the about 0.2– 0.3% DHA found in most LCPUFA- enriched formulae for term infants currently marketed around the world, which, however, generally also contain preformed ARA at levels equal to or often 2-fold higher than the DHA content. The proposed obligatory inclusion of DHA in all infant and follow-on formulae is welcomed by many scientists and pediatricians in view of the indications for beneficial effects [34], but the advice to provide infant formula from birth that supplies DHA but no ARA has been heavily criticized [50]. During pregnancy and infancy, both DHA and ARA are deposited in relatively large amounts in human tissues, including the brain [51, 52]. Fetal accretion of both DHA and ARA during pregnancy is facilitated by their active and preferential maternofetal placental transfer [53]. Pregnant women’s red blood cell levels of both DHA and ARA were positively associated with their children’s IQ at school age [54]. At birth, higher cord blood contents of both DHA and ARA predicted less later behavioral problems, emotional difficulties, hyperactivity, and attention deficit at the age of 10 years [55]. After birth, breastfed infants always get both preformed DHA and ARA, usually with a higher provision of ARA than of DHA [31, 56]. DHA along with ARA have been added to infant formulae since the 1980s in an attempt to approach the nutrient supply and functional effects achieved with breastfeeding [57– 59]. The global Codex Alimentarius standard on the compositional requirements for infant formula stipulates the optional addition of DHA to infant formula, provided that the ARA content is equal to or higher than the DHA content, thus following the model of typical human milk composition [60].
The advice to provide infant formula from birth that supplies DHA but no ARA has been heavily criticized
Infant formulae providing both DHA and ARA have been evaluated in many controlled trials in infants [34]. In contrast, the proposed composition of term infant formula with up to 1% DHA and no ARA is a novel approach that has not been systematically tested for its suitability and safety in healthy infants born at term. ARA is an essential component of all cell membranes. The amount of ARA incorporated into the developing brain during infancy exceeds the deposition of DHA. Although humans can synthesize ARA to some extent from linoleic acid, infants fed formula without preformed ARA tend to develop lower ARA levels in blood plasma and erythrocytes than breastfed infants who receive both DHA and ARA [51, 57, 61]. In preterm infants, provision of high amounts of omega-3 LC-PUFA without a concomitant supply of ARA has been associated with adverse effects on growth [62, 63]. Further concerns regarding the effects of a high supply of DHA without increasing ARA intakes on infants are raised by the findings of a randomized controlled trial assigning term infants to formula providing either no LC-PUFA or different levels of 0.32, 0.64, and 0.96% DHA at the same ARA level of 0.64% [64]. The investigators performed developmental testing of the participating children up to the age of 6 years. Positive effects in tests on word production, a card sorting task, and an intelligence test were observed with the lower DHA dose. However, performance of children assigned to the highest DHA dose of 0.96% but with a reduced ratio of dietary ARA to DHA was attenuated in the MBCDI Word Production Test and the Dimensional Change Card Sort Test at the highest DHA level, and it was attenuated at the two highest DHA levels in the Peabody Picture Vocabulary Test [64]. Thus, in contrast to what might have been expected, an increase of formula DHA contents above 0.32% did not further improve or at least stabilize developmental outcomes, but actually had adverse effects which might well be due to the reduced dietary ARA to DHA ratios provided with the higher DHA levels.
The effects of equivalent formulae with similar DHA and ARA contents on brain composition were tested in infant baboons. Brain composition in various regions was analyzed. The formula with about 1% DHA induced a trend to lower ARA levels in the retina and all the 8 regions of the brain analyzed, with significantly reduced ARA values in the globus pallidus and the superior colliculus, even though the formula contained 0.64% ARA. These observations raise serious concerns that infant formula with high contents of DHA but lack of ARA may induce adverse effects on brain composition and related functional outcomes.
These findings in human infants and in nonhuman primates question the suitability and safety of the compositional requirements stipulated by the new European legislation, i.e. to provide infant formula from birth with up to 1% of fatty acids as DHA without a proportional increase in the intake of ARA. It is generally agreed upon that any major change in infant formula composition should be subjected to a full preclinical and clinical evaluation of nutritional adequacy and safety prior to the wide use and market introduction of such a modified formula [65–70]. Therefore, it appears to be inappropriate and premature to market formula for term infants from birth with 20–50 mg/100 kcal DHA without added ARA in the absence of accountable data on the suitability and safety from a thorough clinical evaluation of this novel approach [50].
It appears to be inappropriate and premature to market formula for term infants from birth with 20–50 mg/ 100 kcal DHA without added ARA
Conclusion
In addition to meeting the infant needs for energy and essential vitamins and PUFA, human milk lipids provide a mixture of MFGM, complex lipids, and bioactive compounds that may have important biological roles in the breastfed infant, for example with regard to the development of nervous and immune functions. Further studies defining the specific components responsible for such effects and the underlying mechanisms could help to design the best options of nutritional interventions. Methodological progress in the field of metabolomics and lipidomics using liquid chromatography coupled with triple mass spectrometry now allows to determine detailed profiles of molecular species of complex lipids in milk as well as in extremely small sample volumes of infant serum or plasma (e.g. 10 μL) with high quantitative precision [71– 74]. Such lipidomic measurements can serve to provide markers for tissue composition [75] and were shown to be associated with important clinical endpoints in children and adults [76–78] . It is therefore likely that the use of these sophisticated and detailed analytical methods, if combined with appropriate bioinformatics strategies, provide the opportunity to obtain better insights into the physiological roles of complex lipids in early life, which may lead to further improvements in nutritional strategies. Progress in biotechnology and food technology offers new avenues for preparing lipid components that can more closely mimic the complex lipid body provided with breastfeeding. Careful exploration and evaluation of the short- and long-term impact in infants could potentially lead to implementation of major improvements for the feeding of infants who cannot be breastfed. Opportunity also exists in improving our understanding of the optimal supply of LC-PUFA in early and later infancy and in the underlying mechanisms and mediators of their effects, e.g. on neurodevelopment and behavior, immune-related health outcomes, such as allergy and asthma, and pulmonary function.
Acknowledgments
The work of the author is financially supported in part by the Commission of the European Community, the 7th Framework Programme Early Nutrition (FP7-289346), the Horizon 2020 Research and Innovation Programme DYNAHEALTH (No 633595), and the European Research Council Advanced Grant METAGROWTH (ERC-2012-AdG, No. 322605). This manuscript does not necessarily reflect the views of the Commission and in no way anticipates the future policy in this area.
Disclosure Statement
The author declares that no financial or other conflict of interest exists in relation to the contents of this article. The production of this paper has been supported by a grant provided by the Nestlé Nutrition Institute.
References
Koletzko B, Agostoni C, Bergmann R, Ritzenthaler K, Shamir R: Physiological aspects of human milk lipids and implications for infant feeding: a workshop report. Acta Paediatr 2011; 100: 1405–1415.
Delplanque B, Gibson R, Koletzko B, Lapillonne A, Strandvik B: Lipid quality in infant nutrition: current knowledge and future opportunities. J Pediatr Gastroenterol Nutr 2015; 61: 8–17.
Krohn K, Demmelmair H, Koletzko B: Macronutrient requirements for growth: fats and fatty acids; in Duggan C, Watkins JB, Koletzko B, Walker WA (eds): Nutrition in Pediatrics, ed 5. Raleigh, People’s Medical Publishing House, 2016, in press.
Grote V, Verduci E, Scaglioni S, Vecchi F, Contarini G, Giovannini M, et al: Breast milk composition and infant nutrient intakes during the first 12 months of life. Eur J Clin Nutr 2016; 70: 250–256.
Michaelsen KF, Skafte L, Badsberg JH, Jorgensen M: Variation in macronutrients in human bank milk: influencing factors and implications for human milk banking. J Pediatr Gastroenterol Nutr 1990; 11: 229–239.
Koletzko B, Rodriguez-Palmero M, Demmelmair H, Fidler N, Jensen R, Sauerwald T: Physiological aspects of human milk lipids. Early Hum Dev 2001; 65(suppl):S3–S18.
Michaelsen KF, Larsen PS, Thomsen BL, Samuelson G: The Copenhagen Cohort Study on Infant Nutrition and Growth: breast-milk intake, human milk macronutrient content, and influencing factors. Am J Clin Nutr 1994; 59: 600–611.
Keating EM, Curtis BA, Slusher TM: Maternal milk volume and breast milk expression: implications for diet and nutrition in infants; in Zibadi S, Watson RR, Preedy VR (eds): Handbook of Dietary and Nutritional Aspects of Human Breast Milk. Wageningen, Wageningen Academic Publishers, 2013, pp 193–213.
Hernell O, Timby N, Domellof M, Lonnerdal B: Clinical benefits of milk fat globule membranes for infants and children. J Pediatr 2016; 173(suppl):S60–S65.
Giuffrida F, Cruz-Hernandez C, Fluck B, Tavazzi I, Thakkar SK, Destaillats F, et al: Quantification of phospholipids classes in human milk. Lipids 2013; 48: 1051–1058.
Tanaka K, Hosozawa M, Kudo N, Yoshikawa N, Hisata K, Shoji H, et al: The pilot study: sphingomyelin-fortified milk has a positive association with the neurobehavioural development of very low birth weight infants during infancy, randomized control trial. Brain Dev 2013; 35: 45–52.
Gurnida DA, Rowan AM, Idjradinata P, Muchtadi D, Sekarwana N: Association of complex lipids containing gangliosides with cognitive development of 6-month-old infants. Early Hum Dev 2012; 88: 595–601.
Zavaleta N, Kvistgaard AS, Graverholt G, Respicio G, Guija H, Valencia N, et al: Efficacy of an MFGM-enriched complementary food in diarrhea, anemia, and micronutrient status in infants. J Pediatr Gastroenterol Nutr 2011; 53: 561–568.
Veereman-Wauters G, Staelens S, Rombaut R, Dewettinck K, Deboutte D, Brummer RJ, et al: Milk fat globule membrane (INPULSE) enriched formula milk decreases febrile episodes and may improve behavioral regulation in young children. Nutrition 2012; 28: 749–752.
Timby N, Hernell O, Vaarala O, Melin M, Lonnerdal B, Domellof M: Infections in infants fed formula supplemented with bovine milk fat globule membranes. J Pediatr Gastroenterol Nutr 2015; 60: 384–389.
Timby N, Domellof E, Hernell O, Lonnerdal B, Domellof M: Neurodevelopment, nutrition, and growth until 12 mo of age in infants fed a low-energy, low-protein formula supplemented with bovine milk fat globule membranes: a randomized controlled trial. Am J Clin Nutr 2014; 99: 860–868.
Mutemberezi V, Guillemot-Legris O, Muccioli GG: Oxysterols: From cholesterol metabolites to key mediators. Prog Lipid Res 2016; 64: 152–169.
Kinney HC, Karthigasan J, Borenshteyn NI, Flax JD, Kirschner DA: Myelination in the developing human brain: biochemical correlates. Neurochem Res 1994; 19: 983–996.
Cartocci V, Servadio M, Trezza V, Pallottini V: Can cholesterol metabolism modulation affect brain function and behavior? J Cell Physiol 2016; 232: 281–286.
Shamir R, Nganga A, Berkowitz D, Diamond E, Lischinsky S, Lombardo D, et al: Serum levels of bile salt-stimulated lipase and breast feeding. J Pediatr Endocrinol Metab 2003; 16: 1289–1294.
Wong WW, Hachey DL, Insull W, Opekun AR, Klein PD: Effect of dietary cholesterol on cholesterol synthesis in breast-fed and formula-fed infants. J Lipid Res 1993; 34: 1403–1411.
Devlin AM, Innis SM, Shukin R, Rioux MF: Early diet influences hepatic hydroxymethyl glutaryl coenzyme A reductase and 7alphahydroxylase mRNA but not low-density lipoprotein receptor mRNA during development. Metabolism 1998; 47: 20–26.
Cruz ML, Wong WW, Mimouni F, Hachey DL, Setchell KD, Klein PD, et al: Effects of infant nutrition on cholesterol synthesis rates. Pediatr Res 1994; 35: 135–140.
Owen CG, Whincup PH, Odoki K, Gilg JA, Cook DG: Infant feeding and blood cholesterol: a study in adolescents and a systematic review. Pediatrics 2002; 110: 597–608.
Owen CG, Whincup PH, Kaye SJ, Martin RM, Davey Smith G, Cook DG, et al: Does initial breastfeeding lead to lower blood cholesterol in adult life? A quantitative review of the evidence. Am J Clin Nutr 2008; 88: 305– 314.
Ip S, Chung M, Raman G, Chew P, Magula N, DeVine D, Trikalinos T, Lau J: Breastfeeding and maternal and infant health outcomes in developed countries. Evid Rep Technol Assess (Full Rep) 2007;(153):1–186 .
ESPGHAN Committee on Nutrition, Agostoni C, Braegger C, Decsi T, Kolacek S, Koletzko B, et al: Breast-feeding: a commentary by the ESPGHAN Committee on Nutrition. J Pediatr Gastroenterol Nutr 2009; 49: 112– 125.
Carnielli VP, Luijendijk IH, Van Goudoever JB, Sulkers EJ, Boerlage AA, Degenhart HJ, et al: Structural position and amount of palmitic acid in infant formulas: effects on fat, fatty acid, and mineral balance. J Pediatr Gastroenterol Nutr 1996; 23: 553–560.
Ailhaud G, Massiera F, Weill P, Legrand P, Alessandri JM, Guesnet P: Temporal changes in dietary fats: role of n-6 polyunsaturated fatty acids in excessive adipose tissue development and relationship to obesity. Prog Lipid Res 2006; 45: 203–236.
Demmelmair H, Baumheuer M, Koletzko B, Dokoupil K, Kratl G: Metabolism of U13Clabeled linoleic acid in lactating women. J Lipid Res 1998; 39: 1389–1396.
Brenna JT, Varamini B, Jensen RG, Diersen- Schade DA, Boettcher JA, Arterburn LM: Docosahexaenoic and arachidonic acid concentrations in human breast milk worldwide. Am J Clin Nutr 2007; 85: 1457–1464.
Del Prado M, Villalpando S, Elizondo A, Rodriguez M, Demmelmair H, Koletzko B: Contribution of dietary and newly formed arachidonic acid to human milk lipids in women eating a low-fat diet. Am J Clin Nutr 2001; 74: 242–247.
Fidler N, Sauerwald T, Pohl A, Demmelmair H, Koletzko B: Docosahexaenoic acid transfer into human milk after dietary supplementation: a randomized clinical trial. J Lipid Res 2000; 41: 1376–1383.
Koletzko B, Boey CCM, Campoy C, Carlson SE, Chang N, Guillermo-Tuazon MA, et al: Current information and Asian perspectives on long-chain polyunsaturated fatty acids in pregnancy, lactation and infancy. Systematic review and practice recommendations from an Early Nutrition Academy workshop. Ann Nutr Metab 2014; 65: 49–80.
Novak EM, Dyer RA, Innis SM: High dietary omega-6 fatty acids contribute to reduced docosahexaenoic acid in the developing brain and inhibit secondary neurite growth. Brain Res 2008; 1237: 136–145.
EFSA Panel on Dietetic Products, Nutrition and Allergies: DHA and ARA and visual development – scientific substantiation of a health claim related to docosahexaenoic acid (DHA) and arachidonic acid (ARA) and visual development pursuant to Article14 of Regulation (EC) No 1924/2006[1]. EFSA J 2009; 941: 1–14.
Qawasmi A, Landeros-Weisenberger A, Bloch MH: Meta-analysis of LCPUFA supplementation of infant formula and visual acuity. Pediatrics 2013; 131:e262–e272.
Qawasmi A, Landeros-Weisenberger A, Leckman JF, Bloch MH: Meta-analysis of long-chain polyunsaturated fatty acid supplementation of formula and infant cognition. Pediatrics 2012; 129: 1141–1149.
Glaser C, Heinrich J, Koletzko B: Role of FADS1 and FADS2 polymorphisms in polyunsaturated fatty acid metabolism. Metabolism 2010; 59: 993–999.
Glaser C, Lattka E, Rzehak P, Steer C, Koletzko B: Genetic variation in polyunsaturated fatty acid metabolism and its potential relevance for human development and health. Matern Child Nutr 2011; 7(suppl 2):27–40.
Lattka E, Illig T, Koletzko B, Heinrich J: Genetic variants of the FADS1 FADS2 gene cluster as related to essential fatty acid metabolism. Curr Opin Lipidol 2010; 21: 64–69.
Lattka E, Rzehak P, Szabo E, Jakobik V, Weck M, Weyermann M, et al: Genetic variants in the FADS gene cluster are associated with arachidonic acid concentrations of human breast milk at 1.5 and 6 mo postpartum and influence the course of milk dodecanoic, tetracosenoic, and trans-9-octadecenoic acid concentrations over the duration of lactation. Am J Clin Nutr 2011; 93: 382–391.
Steer CD, Davey Smith G, Emmett PM, Hibbeln JR, Golding J: FADS2 polymorphisms modify the effect of breastfeeding on child IQ. PLoS One 2010; 5:e11570.
Morales E, Bustamante M, Gonzalez JR, Guxens M, Torrent M, Mendez M, et al: Genetic variants of the FADS gene cluster and ELOVL gene family, colostrums LC-PUFA levels, breastfeeding, and child cognition. PLoS One 2011; 6:e17181.
Jensen CL, Maude M, Anderson RE, Heird WC: Effect of docosahexaenoic acid supplementation of lactating women on the fatty acid composition of breast milk lipids and maternal and infant plasma phospholipids. Am J Clin Nutr 2000; 71(1 suppl):292S–9S.
Standl M, Sausenthaler S, Lattka E, Koletzko S, Bauer CP, Wichmann HE, et al: FADS gene cluster modulates the effect of breastfeeding on asthma. Results from the GINIplus and LISAplus studies. Allergy 2012; 67: 83–90.
European Commission: Commission Delegated Regulation (EU) 2016/127 of 25 September 2015 supplementing Regulation (EU) No 609/2013 of the European Parliament and of the Council as regards the specific compositional and information requirements for infant formula and follow-on formula and as regards requirements on information relating to infant and young child feeding. Official Journal of the European Union 2016:L25/1.
EFSA Panel on Dietetic Products, Nutrition and Allergies: Scientific opinion on nutrient requirements and dietary intakes of infants and young children in the European Union. EFSA J 2013; 11: 3408.
EFSA Panel on Dietetic Products, Nutrition and Allergies: Scientific opinion on the essential composition of infant and follow-on formulae. EFSA J 2014; 12: 106.
Koletzko B, Carlson SE, van Goudoever JB: Should infant formula provide both omega-3 DHA and omega-6 arachidonic acid? Ann Nutr Metab 2015; 66: 137–138.
Makrides M, Neumann MA, Byard RW, Simmer K, Gibson RA: Fatty acid composition of brain, retina, and erythrocytes in breast- and formula-fed infants. Am J Clin Nutr 1994; 60: 189–194.
Martinez M: Tissue levels of polyunsaturated fatty acids during early human development. J Pediatr 1992; 120:S129–S138.
Larque E, Ruiz-Palacios M, Koletzko B: Placental regulation of fetal nutrient supply. Curr Opin Clin Nutr Metab Care 2013; 16: 292–297.
Steer CD, Lattka E, Koletzko B, Golding J, Hibbeln JR: Maternal fatty acids in pregnancy, FADS polymorphisms, and child intelligence quotient at 8 y of age. Am J Clin Nutr 2013; 98: 1575–1582.
Kohlboeck G, Glaser C, Tiesler C, Demmelmair H, Standl M, Romanos M, et al: Effect of fatty acid status in cord blood serum on children’s behavioral difficulties at 10 y of age: results from the LISAplus Study. Am J Clin Nutr 2011; 94: 1592–1599.
Demmelmair H, Koletzko B: Importance of fatty acids in the perinatal period. World Rev Nutr Diet 2015; 112: 31–47.
Koletzko B, Schmidt E, Bremer HJ, Haug M, Harzer G: Effects of dietary long-chain polyunsaturated fatty acids on the essential fatty acid status of premature infants. Eur J Pediatr 1989; 148: 669–675.
Carlson SE, Cooke RJ, Rhodes PG, Peeples JM, Werkman SH: Effect of vegetable and marine oils in preterm infant formulas on blood arachidonic and docosahexaenoic acids. J Pediatr 1992; 120:S159–S167.
Makrides M, Neumann MA, Simmer K, Gibson RA: Erythrocyte fatty acids of term infants fed either breast milk, standard formula, or formula supplemented with long-chain polyunsaturates. Lipids 1995; 30: 941–948.
Codex Alimentarius Commission: Standard for infant formula and formulas for special medical purposes intended for infants. Codex Stan 72 – 1981. Rome, Codex Alimentarius Commission 2007, pp 1–21.
Koletzko B, Sauerwald T, Demmelmair H, Herzog M, von Schenck U, Bohles H, et al: Dietary long-chain polyunsaturated fatty acid supplementation in infants with phenylketonuria: a randomized controlled trial. J Inherit Metab Dis 2007; 30: 326–332.
Carlson SE, Werkman SH, Peeples JM, Cooke RJ, Tolley EA: Arachidonic acid status correlates with first year growth in preterm infants. Proc Natl Acad Sci USA 1993; 90: 1073–1077.
Carlson SE, Cooke RJ, Werkman SH, Tolley EA: First year growth of preterm infants fed standard compared to marine oil n-3 supplemented formula. Lipids 1992; 27: 901–907.
Colombo J, Carlson SE, Cheatham CL, Shaddy DJ, Kerling EH, Thodosoff JM, et al: Long-term effects of LCPUFA supplementation on childhood cognitive outcomes. Am J Clin Nutr 2013; 98: 403–412.
Committee on the Evaluation of the Addition of Ingredients New to Infant Formula, Food and Nutrition Board, Institute of Medicine: Infant Formula. Evaluating the Safety of New Ingredients. Washington, The National Academy Press, 2001.
ESPGHAN Committee on Nutrition, Aggett PJ, Agostini C, Goulet O, Hernell O, Koletzko B, et al: The nutritional and safety assessment of breast milk substitutes and other dietary products for infants: a commentary by the ESPGHAN Committee on Nutrition. J Pediatr Gastroenterol Nutr 2001; 32: 256– 258.
Koletzko B, Ashwell M, Beck B, Bronner A, Mathioudakis B: Characterisation of infant food modifications in the European Union. Ann Nutr Metab 2002; 46: 231–242.
Koletzko B, Szajewska H, Ashwell M, Shamir R, Aggett P, Baerlocher K, et al: Documentation of functional and clinical effects of infant nutrition: setting the scene for COMMENT. Ann Nutr Metab 2012; 60: 222–232.
Koletzko B, Shamir R, Ashwell M: Quality and safety aspects of infant nutrition. Ann Nutr Metab 2012; 60: 179–184.
European Commission, Scientific Committee on Food; Koletzko B, Saris WH, Flynn A, Palou A, Wal JM, et al: Report of the Scientific Committee on Food on the Revision of Essential Requirements of Infant Formulae and Follow-on Formulae. Brussels, European Commission, 2003.
Uhl O, Hellmuth C, Demmelmair H, Zhou SJ, Makrides M, Prosser C, et al: Dietary effects on plasma glycerophospholipids. J Pediatr Gastroenterol Nutr 2015; 61: 367–372.
Uhl O, Glaser C, Demmelmair H, Koletzko B: Reversed phase LC/MS/MS method for targeted quantification of glycerophospholipid molecular species in plasma. J Chromatogr B Analyt Technol Biomed Life Sci 2011; 879: 3556–3564.
Hellmuth C, Uhl O, Segura-Moreno M, Demmelmair H, Koletzko B: Determination of acylglycerols from biological samples with chromatography-based methods. J Sep Sci 2011; 34: 3470–3483.
Hellmuth C, Weber M, Koletzko B, Peissner W: Nonesterified fatty acid determination for functional lipidomics: comprehensive ultrahigh performance liquid chromatography- tandem mass spectrometry quantitation, qualification, and parameter prediction. Anal Chem 2012; 84: 1483–1490.
Hellmuth C, Demmelmair H, Schmitt I, Peissner W, Bluher M, Koletzko B: Association between plasma nonesterified fatty acids species and adipose tissue fatty acid composition. PLoS One 2013; 8:e74927.
Rauschert S, Uhl O, Koletzko B, Kirchberg F, Mori TA, Huang RC, et al: Lipidomics reveals associations of phospholipids with obesity and insulin resistance in young adults. J Clin Endocrinol Metab 2016; 101: 871–879.
Reinehr T, Wolters B, Knop C, Lass N, Hellmuth C, Harder U, et al: Changes in the serum metabolite profile in obese children with weight loss. Eur J Nutr 2015; 54: 173–181.
Rzehak P, Hellmuth C, Uhl O, Kirchberg FF, Peissner W, Harder U, et al: Rapid growth and childhood obesity are strongly associated with LysoPC(14: 0). Ann Nutr Metab 2014; 64: 294–303.
Khan S, Hepworth AR, Prime DK, Lai CT, Trengove NJ, Hartmann PE: Variation in fat, lactose, and protein composition in breast milk over 24 hours: associations with infant feeding patterns. J Hum Lact 2013; 29: 81–89.